ГДЗ Геометрия 9 класс Мерзляк, Полонский, Якир
Геометрия 9 класс
Учебник
Мерзляк, Полонский, Якир
Алгоритм успеха
Вентана-Граф
Нюансы школьной программы не позволяют относиться к ней походя, не уделяя ей достаточного внимания. Порой возникает ощущение, что сама подача материала намеренно усложняется авторами учебных пособий, которые стараются полностью соответствовать ФГОСу. Подросткам бывает иногда трудно понять что же именно обозначает то или иное высказывание и как именно это соотносится с практическими заданиями. Учителя же не стремятся сразу внести ясность в такое положение вещей. Поэтому решебник к учебнику «Геометрия 9 класс» Мерзляк, Полонский, Якир является универсальным средством, где обширно отражена вся информация.
Основные моменты данного сборника
В пособии имеется восемьсот восемьдесят шесть упражнений, которые распределены по тематическим разделам. Так же авторы предоставили доскональные ответы к вопросам по параграфам и к тестовым заданиям. ГДЗ по геометрии 9 класс Мерзляк не раз пригодится учащимся в этом году как дополнительное средство по разъяснению материала.
ГДЗ по геометрии 9 класс Мерзляк не раз пригодится учащимся в этом году как дополнительное средство по разъяснению материала.
Почему решебник необходимо использовать
Для некоторых ребят после окончания этого класса уже начнется совершенно другая жизнь. Впереди их ждет колледж, институт, работа, а пока что необходимо приложить побольше усилий, чтобы успешно преодолеть все препятствия этого курса. А их будет немало, так же как и той информации, которую нужно будет усвоить. А так как уже достаточно давно появилась тенденция, что школьники должны сами разбираться в новом материале, то и в этот раз все ложиться на их плечи. Однако существует много аспектов, разобраться самостоятельно в которых просто невозможно. Поэтому решебник к учебнику «Геометрия 9 класс» Мерзляк может оказаться весьма кстати, когда необходимо разобрать то или иное упражнение, чтобы понять принцип его выполнения. «Вентана-граф», 2016 г.
ГДЗ по Геометрии для 9 класса А.
 Г. Мерзляк, В.Б. Полонский, М.С. Якир на 5
Г. Мерзляк, В.Б. Полонский, М.С. Якир на 5
Авторы: А.Г. Мерзляк, В.Б. Полонский, М.С. Якир.
Издательство:
Вентана-граф 2016
В девятом классе ребята на уроке геометрии будут уверенно справляться со всеми трудностями, если обратятся за удалёнными консультациями к содержательному интернет-самоучителю – «ГДЗ по Геометрии 9 класс Мерзляк, Полонский (Вентана-граф)». Геометрия является важным элементом общеобразовательного обучения. Рассматривает объекты в пространстве, их свойства, взаимодействие, а также уникальные алгоритмы доказательств теорем, различные аксиомы, известные науке.
Приобретённые знания и умения на уроках в первую очередь пригодятся будущим инженерам, архитекторам, а также художникам. В обыденной жизни навыки распознавания различных геометрических форм позволяют лучше ориентироваться на местности, демонстрируют высокий уровень образованности и интеллекта человека.
Рабочая программа по геометрии
Девятиклассники на занятиях технической дисциплины будут рассматривать весьма непростые темы, с которыми может не справиться даже самый сообразительный подросток. Разобрать понятия и определения особенно трудно без лекций преподавателя. Зачастую случается пропускать уроки по вынужденным причинам. В этом случае поможет ГДЗ. Он с лёгкостью наверстает вместе с учеником все пробелы, чтобы проблемы не копились, как «снежный ком».
Рассмотрим самые затруднительные параграфы из содержания учебно-методического комплекта по геометрии:
- – уравнение прямой;
- – как звучит теорема косинусов;
- – скалярное произведение векторов.
«ГДЗ по Геометрии за 9 класс Мерзляк А. Г., Полонский В. Б., Якир М. С. (Вентана-граф)» объяснит любой непонятный момент из учебника. Геометрия на поверку окажется не такой сложной, как казалась ранее.
Чем хорош решебник по геометрии для 9 класса от Мерзляка
Главная цель справочника – упростить образовательный процесс по геометрии. Ученикам удастся справляться с домашними заданиями оперативно, без лишних переживаний. При этом в голове останется больше полезной информации, так как в ходе самопроверки по решебнику придётся ещё раз повторить изученное. Прочие плюсы образовательного процесса под руководством ГДЗ:
Ученикам удастся справляться с домашними заданиями оперативно, без лишних переживаний. При этом в голове останется больше полезной информации, так как в ходе самопроверки по решебнику придётся ещё раз повторить изученное. Прочие плюсы образовательного процесса под руководством ГДЗ:
- – онлайн-формат;
- – доступен со смартфона;
- – к некоторым особо сложным номерам упражнений присутствуют уточняющие комментарии для лучшего разбора.
Сборник верных ответов досконально проверен специалистами в данной технической области, соответствует федеральному государственному образовательному стандарту.
Решебник ⏩ ГДЗ Геометрия 9 класс ⚡ А. Г. Мерзляк, В. Б. Полонский, М. С. Якир 2009
ГДЗ геометрия 9 класс Мерзляк
Каждый в школе думал о том, когда же уже закончится этот урок геометрии или алгебры. Вряд ли найдутся школьники, которые с нетерпением ждут этих уроков. Как ни крути, а геометрия необходимая наука, которая еще несколько тысяч лет назад вызывала интерес и изучение.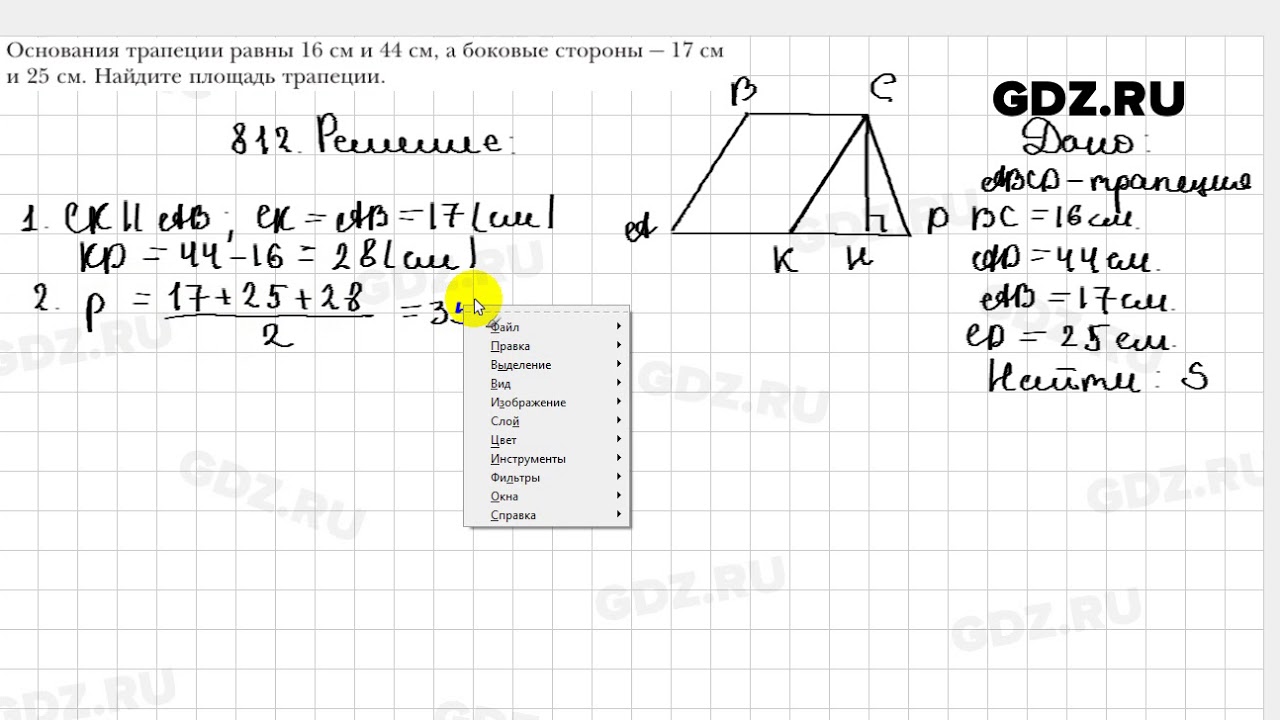 Без геометрии не высчитать нужных углов, площадь и многие другие вещи, необходимые при строительстве или обработке земли. Да, скорее всего многие изучаемые теоремы и формулы Вам не понадобятся, но они необходимы, как общая основа школьных знаний. Если Вам сложно дается геометрия, и необходимо подтянуть оценки, но помочь в этом деле некому, тогда лучшим вариантом станет ГДЗ геометрия 9 класс Мерзляк. В этом решебнике имеются ответы на все задания из школьного учебника, что позволит больше не испытывать проблем во время выполнения домашней или самостоятельной работы.
Без геометрии не высчитать нужных углов, площадь и многие другие вещи, необходимые при строительстве или обработке земли. Да, скорее всего многие изучаемые теоремы и формулы Вам не понадобятся, но они необходимы, как общая основа школьных знаний. Если Вам сложно дается геометрия, и необходимо подтянуть оценки, но помочь в этом деле некому, тогда лучшим вариантом станет ГДЗ геометрия 9 класс Мерзляк. В этом решебнике имеются ответы на все задания из школьного учебника, что позволит больше не испытывать проблем во время выполнения домашней или самостоятельной работы.
Решебник геометрия 9 класс Мерзляк онлайн или скачать
Решебник удобно разделен на главы и параграфы, что обеспечивает быстрое нахождение нужных ответов. К тому же, все ответы в наших ГДЗ детально расписанные, и это не для того, чтобы учитель не сомневался в том, что Вы сделали задание самостоятельно. Расписанные задания дают Вам возможность разобраться в задачах самостоятельно, увидеть каким способом они были решены и какие формулы при этом использовались.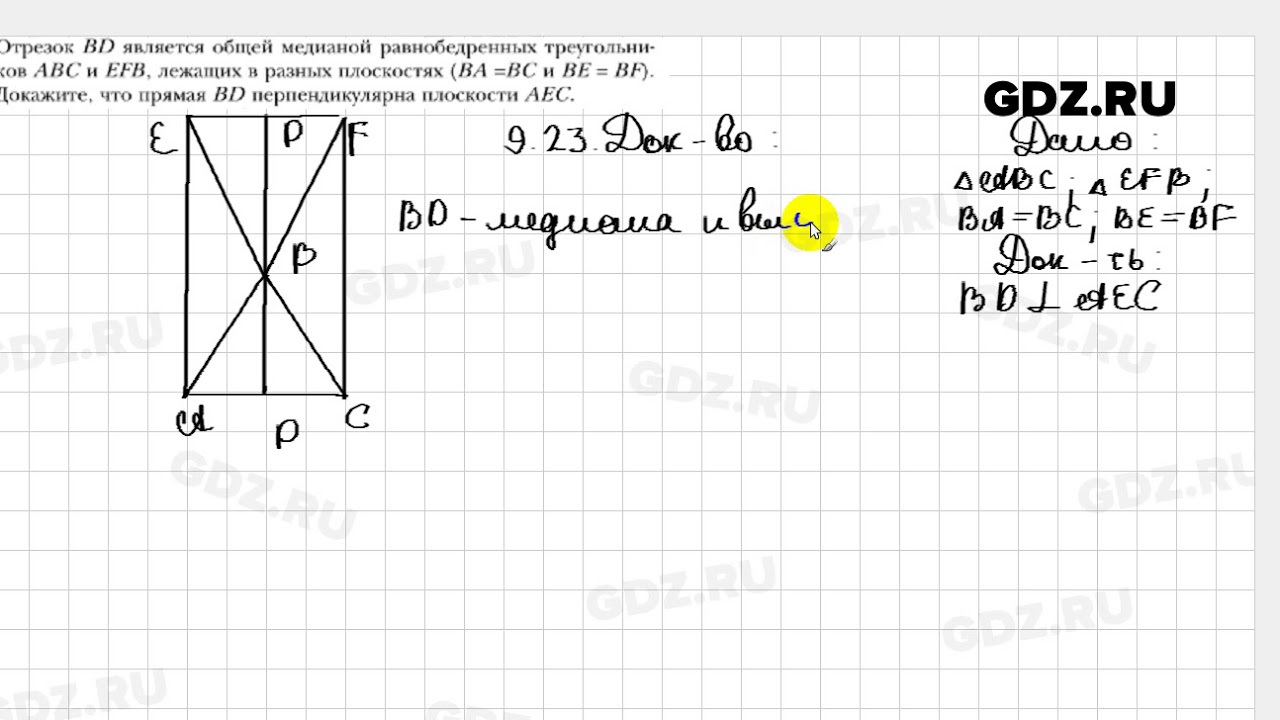 То есть, Вы можете, например, сначала попытаться сделать задание самостоятельно, а уже после – посмотреть правильный ответ. Таким образом, понемногу Вы начнете понимать материал и изучаемую тему.
То есть, Вы можете, например, сначала попытаться сделать задание самостоятельно, а уже после – посмотреть правильный ответ. Таким образом, понемногу Вы начнете понимать материал и изучаемую тему.
Сегодня, имеются много различных видов учебников по геометрии, поэтому, смотрите внимательно фамилию автора перед загрузкой решебника. Наша команда старается обновлять ГДЗ по мере необходимости, вот почему, скачав его, Вы можете не сомневаться, что в нем найдется нужный ответ. К тому же наши ГДЗ включают в себя не только часть ответов, а полностью все, что делает их лучшими среди аналогов. Загружайте решебник геометрия 9 класс Мерзляк и забудьте навсегда о проблемах и переживаниях, которые когда-то у Вас вызывал этот предмет. Загружайте ГДЗ с нашего сайта прямо сейчас, и убедитесь сами в его удобности и практичности.
Введение
Введение
Циклическая перестановка GFP и его вариантов заметно расширила возможности использования флуоресцентных белков (FP), позволив разработать генетически кодируемые сенсоры [1], [2], [3] и облегчив использование FP во флуоресценции. комплементарные анализы [4], [5]. Распространение этой технологии на красные флуоресцентные белки, хотя и потенциально весьма полезное, было ограничено; яркие и термостабильные кольцевые пермутанты или комплементарные пептиды было трудно разработать [1], [2], вероятно, потому, что локальное окружение хромофора изменяется в результате перегруппировок, предпринятых до сих пор [6].Стремясь устранить это ограничение, мы предприняли систематические усилия для получения ярких и стабильных циклически пермутированных вариантов двух встречающихся в природе белков, mCherry, мономерного варианта Discosoma sp. коралловый белок DsRed [7] и mKate, мономерный вариант белка eqFP578 анемоны Entacmaea quadricolor [8], [9]. Оба белка обладают полезными свойствами визуализации тканей, включая характеристики относительной яркости и длинноволнового излучения; более того, оба они спектрально отличаются от GFP и его производных, потенциально расширяя цветовую палитру генетически кодируемых сенсоров или комплементарных пар, которые можно использовать одновременно с конструкциями на основе GFP.
комплементарные анализы [4], [5]. Распространение этой технологии на красные флуоресцентные белки, хотя и потенциально весьма полезное, было ограничено; яркие и термостабильные кольцевые пермутанты или комплементарные пептиды было трудно разработать [1], [2], вероятно, потому, что локальное окружение хромофора изменяется в результате перегруппировок, предпринятых до сих пор [6].Стремясь устранить это ограничение, мы предприняли систематические усилия для получения ярких и стабильных циклически пермутированных вариантов двух встречающихся в природе белков, mCherry, мономерного варианта Discosoma sp. коралловый белок DsRed [7] и mKate, мономерный вариант белка eqFP578 анемоны Entacmaea quadricolor [8], [9]. Оба белка обладают полезными свойствами визуализации тканей, включая характеристики относительной яркости и длинноволнового излучения; более того, оба они спектрально отличаются от GFP и его производных, потенциально расширяя цветовую палитру генетически кодируемых сенсоров или комплементарных пар, которые можно использовать одновременно с конструкциями на основе GFP. Здесь мы сообщаем о систематической оценке сайтов круговой перестановки в mCherry и mKate, а также о разработке высокоэффективных красных флуоресцентных белков с круговой перестановкой (cp-RFP).
Здесь мы сообщаем о систематической оценке сайтов круговой перестановки в mCherry и mKate, а также о разработке высокоэффективных красных флуоресцентных белков с круговой перестановкой (cp-RFP).
Материалы и методы. Генерация cp-RFP из шаблонов тандемного слияния
. Чтобы эффективно исследовать несколько сайтов перестановок, мы создали шаблоны тандемного слияния [10] mKate (pRSET-tdmKate) и mCherry (pRSET-tdmCherry). Для первого кодирующая последовательность mKate (231 AA) была амплифицирована из плазмиды TagFP635 (Evrogen) с использованием ДНК-полимеразы Phusion High-Fidelity (Finnzymes) с копией 1, прямой (F1) BamHI-mK-1F (atcaggatccatgtctgagctgattaaggaga) и обратной (R1 ) Праймеры HindIII-mK-231R (ctacaagctttcatttgtgccccagttgctagg) (сайты рестрикции выделены курсивом).Амплифицированный продукт встраивали в сайты BamHI/HindIII pRSET-A (Invitrogen) для получения pRSET-mKate. Вторую копию рамки считывания mKate амплифицировали с матрицы TagFP635 с праймерами (F2) BamHI-mK-1F и (R2) BamHI-mK-231R-linker (ctacggatccgccggtaccgccttg tgccccagtttgctagg). Обратите внимание на отсутствие стоп-кодона в обратном праймере (R2) и подчеркнутом линкере, так что сайт BamHI обеспечивает остатки GS в линкерном пептиде GGTGGS. Продукт ПЦР вставляли в сайт BamHI pRSET-mKate, и правильную ориентацию определяли с помощью ПЦР и секвенирования ДНК.Конечная плазмида (pRSET-tdmKate) содержит две тандемные кодирующие последовательности mKate, разделенные последовательностью, кодирующей линкерный пептид GGTGGS, в непрерывной рамке считывания (фиг. 1А). Аналогичная стратегия была использована для конструирования pRSET-tdmCherry с использованием кодирующей последовательности mCherry (236 AA) из pRSET-B mCherry [7], праймеры (F1) BamHI-mC-1F (ctacggatccatggtgagcaagggcgaggagga), (R1) HindIII-mC-236R ( ctacaagctttcacttgtacagctcgtccat), (F2) BamHI-mC-1F и (R2) BamHI-mC-236R-линкер (ctacggatccgccggtaccgcccttgtacagctcgtcca).Матрицы тандемного слияния использовали в качестве матриц ПЦР для создания cp-RFP путем систематического изменения N- и C-концевых праймеров. Для конструирования cp-mKate 156-155 сайт XhoI был заменен на NheI в прямом праймере, а амплифицированный фрагмент был клонирован в сайты NheI и EcoRI pRSET-A, чтобы предотвратить воссоздание cp-mKate 154-155.
Обратите внимание на отсутствие стоп-кодона в обратном праймере (R2) и подчеркнутом линкере, так что сайт BamHI обеспечивает остатки GS в линкерном пептиде GGTGGS. Продукт ПЦР вставляли в сайт BamHI pRSET-mKate, и правильную ориентацию определяли с помощью ПЦР и секвенирования ДНК.Конечная плазмида (pRSET-tdmKate) содержит две тандемные кодирующие последовательности mKate, разделенные последовательностью, кодирующей линкерный пептид GGTGGS, в непрерывной рамке считывания (фиг. 1А). Аналогичная стратегия была использована для конструирования pRSET-tdmCherry с использованием кодирующей последовательности mCherry (236 AA) из pRSET-B mCherry [7], праймеры (F1) BamHI-mC-1F (ctacggatccatggtgagcaagggcgaggagga), (R1) HindIII-mC-236R ( ctacaagctttcacttgtacagctcgtccat), (F2) BamHI-mC-1F и (R2) BamHI-mC-236R-линкер (ctacggatccgccggtaccgcccttgtacagctcgtcca).Матрицы тандемного слияния использовали в качестве матриц ПЦР для создания cp-RFP путем систематического изменения N- и C-концевых праймеров. Для конструирования cp-mKate 156-155 сайт XhoI был заменен на NheI в прямом праймере, а амплифицированный фрагмент был клонирован в сайты NheI и EcoRI pRSET-A, чтобы предотвратить воссоздание cp-mKate 154-155. вариант (рис. 1Б,С). Укороченные варианты cp-mKate были сконструированы из шаблона тандемного слияния mKate с использованием аналогичной стратегии ПЦР.
вариант (рис. 1Б,С). Укороченные варианты cp-mKate были сконструированы из шаблона тандемного слияния mKate с использованием аналогичной стратегии ПЦР.
10.1371/journal.pone.0020505.g001Рисунок 1
A. Циклические пермутанты были сконструированы из матрицы тандемного слияния с прямым (новый N-концевой) и обратным (новый C-концевой) праймерами, давая ампликон полной длины, начинающийся и заканчивающийся в любом желаемом сайте. Показана стратегия круговой перестановки mKate. B. Схема конечных белков cp-RFP с N-концевым лидерным пептидом из вектора экспрессии pRSET-A; C (n+1-231) и N (1-n)-концевые остатки mKate были соединены линкером GGTGGS, а линкер LE между RSET и cp-mKate кодировался сайтом XhoI.C. Аминокислотные последовательности DsRed из Discosoma sp. коралл, mCherry, полученный из DsRed, и mKate из актинии Entacmaea quadricolor. Остатки, образующие хромофоры, подчеркнуты; остатки, образующие бета-цепи, выделены жирным шрифтом.
Скрининг cp-RFP в клетках E.coli
E.coli BL21 Star (DE3) pLysS (Invitrogen) трансформировали вариантными плазмидами pREST-cp-mCherry или pRSET-cp-mKate и высевали на чашки с агаром LB с добавлением 100 мкг /мл ампициллин.После инкубации при 37°C в течение 24 часов колонии подвергали скринингу на предмет красной флуоресценции с использованием широкопольной системы макроизображения (OV100, Olympus, Япония; возбуждение 545 нм/испускание 570–625 нм). Колонии подвергали повторному скринингу через 1–7 дней при 2–8°С. Изображения были проанализированы на яркость с использованием изображения J и нормализованы для колоний, трансформированных mCherry или mKate; контрольные колонии переносчиков использовали в качестве нулевого фона флуоресценции.
Экспрессия и очистка белков
Ночные посевные культуры трансформированных клеток BL21 Star (DE3) pLysS использовали для инокуляции (разведение 1×40) 500 мл бульона Terrific, содержащего 100 мкг/мл ампициллина. Культуры инкубировали при 37°С (250 об/мин) до ОП 600 ~1,0; Добавляли 0,5 мМ ИПТГ и инкубировали 16 ч при 20°С (250 об/мин). Клетки собирали центрифугированием при 6000×g в течение 10 минут при 4°C и ресуспендировали в 20 мМ HEPES (pH 7,4) с 350 мМ NaCl и 0,1% TritonX-100. Культуры обрабатывали ультразвуком и удаляли клеточный дебрис центрифугированием при 20000×g в течение 30 минут при 4°C. Очищенный лизат смешивали с 3 мл Ni-Charged Resins Profinity IMAC (Bio-Rad) и инкубировали на качалке в течение 5 минут, смолу выливали на 0.Хроматографическая колонка 8×4 см (Bio-Rad), промытая 3 раза 20 мМ буфером HEPES (pH 7,4), содержащим 350 мМ NaCl и 0,1% TritonX-100, и 3 раза 20 мМ HEPES (pH 7,4), содержащим 350 мМ NaCl. ; за каждой промывкой следовало низкоскоростное центрифугирование при 800×g в течение 30 секунд. Белки элюировали 20 мМ HEPES (pH 7,8), содержащим 350 мМ NaCl и 300 мМ имидазольного буфера, а объединенные белковые фракции обессоливали и концентрировали с использованием устройства Amicon Ultra-15 (Millipore) с 20 мМ HEPES (pH 7,4), заправка 2–2.
Культуры инкубировали при 37°С (250 об/мин) до ОП 600 ~1,0; Добавляли 0,5 мМ ИПТГ и инкубировали 16 ч при 20°С (250 об/мин). Клетки собирали центрифугированием при 6000×g в течение 10 минут при 4°C и ресуспендировали в 20 мМ HEPES (pH 7,4) с 350 мМ NaCl и 0,1% TritonX-100. Культуры обрабатывали ультразвуком и удаляли клеточный дебрис центрифугированием при 20000×g в течение 30 минут при 4°C. Очищенный лизат смешивали с 3 мл Ni-Charged Resins Profinity IMAC (Bio-Rad) и инкубировали на качалке в течение 5 минут, смолу выливали на 0.Хроматографическая колонка 8×4 см (Bio-Rad), промытая 3 раза 20 мМ буфером HEPES (pH 7,4), содержащим 350 мМ NaCl и 0,1% TritonX-100, и 3 раза 20 мМ HEPES (pH 7,4), содержащим 350 мМ NaCl. ; за каждой промывкой следовало низкоскоростное центрифугирование при 800×g в течение 30 секунд. Белки элюировали 20 мМ HEPES (pH 7,8), содержащим 350 мМ NaCl и 300 мМ имидазольного буфера, а объединенные белковые фракции обессоливали и концентрировали с использованием устройства Amicon Ultra-15 (Millipore) с 20 мМ HEPES (pH 7,4), заправка 2–2. 3 раза.Чистоту белка проверяли с помощью SDS-PAGE и измеряли концентрацию (Pierce BCA Protein Assay).
3 раза.Чистоту белка проверяли с помощью SDS-PAGE и измеряли концентрацию (Pierce BCA Protein Assay).
Спектроскопия cp-RFP
Спектральные свойства RSET-меченых белков в 20 мМ HEPES (pH 7,4) измеряли при 20°C. Спектры поглощения регистрировали на спектрофотометре УФ/видимой области DU 730 (Beckman Coulter), спектры флуоресценции – на спектрофлуориметре FluoroMax-3 (Jobin Yvon Horiba). Коэффициенты экстинкции белков cp-mCherry измеряли методом щелочной денатурации с 0.1 М NaOH [11], [12]. Квантовый выход (Φ) для cp-mCherrys и cp-mKates измеряли с использованием исходных молекул (mCherry: Φ = 0,22; mKate: Φ = 0,33) в качестве стандартов сравнения [13]. Флуоресценцию 5 мкМ белковых растворов в 20 мМ HEPES (рН 7,4) измеряли с помощью многорежимного считывателя микропланшетов Synergy 2 (Biotek; возбуждение 575/15, эмиссия 620/15).
Кристаллизация и определение структуры
Для кристаллизации кодирующая область cp-mKate 154-153 и cp-mKate 168-167 была амплифицирована и клонирована в модифицированную экспрессионную плазмиду pET28a (Novagen) с получением слитого белка SUMO с N-концевым гексагистидином. , с меченым фрагментом, расщепляемым протеазой Ulp-1 из S.шейки матки.
, с меченым фрагментом, расщепляемым протеазой Ulp-1 из S.шейки матки.
клетки E.coli BL21 (DE3) (Novagen) трансформировали плазмидами cp-mKate. Экспрессию и очистку белков проводили по протоколам, описанным ранее [6]. Кристаллы получали диффузией паров висячих капель путем смешивания равных объемов белка (∼35 мг/мл) и резервуарного раствора с последующей инкубацией при 20°С. Кристаллы получали с резервуарным раствором, содержащим 0,1 М Трис-HCl pH 7,4, 0,2 М MgCl 2 и 18% ПЭГ3350 (для cp-mKate 154-153 ) или 0.1 М дигидрат формиата магния и 14% ПЭГ 3350 (для cp-mKate 168-167 ). Все кристаллы были криозащищены с использованием кристаллизационных растворов с добавлением 20% ксилита, заморожены в жидком азоте и выдерживались при 100 К во время сбора данных.
Наборы данных были собраны с использованием синхротронного излучения в Корнеллском синхротронном источнике высоких энергий (CHESS, Итака). Обработку данных проводили с помощью пакета программ HKL2000 [14]. Фазы получали путем молекулярного замещения с помощью пакета программ PHENIX [15] с доступной структурой mKate (1.8 Å, pH 2,0, код PDB: 3BX9) [16] в качестве поисковых моделей. Ручное уточнение в COOT [17] и минимизация с помощью PHENIX [15] дали окончательные модели с хорошей геометрией.
Фазы получали путем молекулярного замещения с помощью пакета программ PHENIX [15] с доступной структурой mKate (1.8 Å, pH 2,0, код PDB: 3BX9) [16] в качестве поисковых моделей. Ручное уточнение в COOT [17] и минимизация с помощью PHENIX [15] дали окончательные модели с хорошей геометрией.
Результаты Циркулярная перестановка mCherry
Для эффективного сканирования mCherry на наличие многообещающих участков круговой перестановки мы создали несколько клонов cp-mCherry с помощью ПЦР-амплификации матрицы тандем-mCherry (pRSET-tdmCherry, методы) и непосредственно сравнили красную флуоресценцию бактериальных колоний с бактериальными колониями, трансформированными PRSET-mCherry.Первоначальные конструкции cp-mCherry нацелены на петли mCherry от 4 th до 10 th β-нитей (рис. 2). Ни одна из колоний варианта cp-mCherry не показала заметной флуоресценции после инкубации в течение ночи при 37°C по сравнению с колониями, трансформированными mCherry дикого типа, тогда как после дальнейшей инкубации в течение 24 ч либо при комнатной температуре, либо при 2–8°C флуоресценция была обнаружена в cp. -mCherry 158-157 , cp-mCherry 175-174 и cp-mCherry 193-192 колоний.Колонии других вариантов не проявляли заметной флуоресценции даже через 7 дней при 2–8°C (данные не представлены).
-mCherry 158-157 , cp-mCherry 175-174 и cp-mCherry 193-192 колоний.Колонии других вариантов не проявляли заметной флуоресценции даже через 7 дней при 2–8°C (данные не представлены).
10.1371/journal.pone.0020505.g002Рисунок 2
mCherry и mKate были пронумерованы по первичным аминокислотным последовательностям, как показано на фиг. 1C. Открытые кружки обозначают участки с флуоресценцией, закрытые кружки — участки без флуоресценции. Три высоко гомологичных участка устойчивых к круговой перестановке сайтов у mCherry и mKate: область петли 7–8, расположенная в петле между 7 th и 8 th β-тяжами и фланкирующими сайтами на β цепях, петля 8–9 область, расположенная в петле между 8 й и 9 й β-тяжами и фланкирующими участками на β тяжах, и область петли 9–10, фланкированная 9 й и 10 й β нитями.
Первоначальный скрининг выявил три сайта в отдельных петлях, соединяющих β-цепи. Мы исследовали фланкирующие последовательности в каждом из этих сайтов, создавая и анализируя дополнительные варианты cp-mCherry. Самая яркая колония (cp-mCherry 194-193 ) демонстрировала только 1% mCherry после 24-часовой инкубации при 37°C, что указывает на медленное созревание циклически пермутированных белков в E.coli. Основываясь на этом наблюдении, все скрининги оценивали через 24 часа при 37°C и еще через 72 часа при 2–8°C (таблица 1).Данные флуоресценции колоний показали, что пермиссивные сайты были сгруппированы в три петли между 7 th и 8 th β-нитями, 8 th и 9 th β-нитями, 9 th и 10 th th th th th th th th 0 тяжей и доходящих до концов фланкирующих β-тяжей в каждой области соответственно (рис. 2). Валидация анализа флуоресценции бактериальных колоний с измерением флуоресценции очищенных белков cp-mCherry в 20 мМ HEPES (pH 7,4) показала, что cp-mCherry 194-193 является самым ярким вариантом из всех проверенных круговых перестановок, проявляя примерно 90% относительная яркость на хромофорной основе и 60. Белковая яркость 6% по сравнению с mCherry (таблица 2). Разница между этими измерениями указывает на субоптимальную укладку белка, как обсуждается ниже. Ранее сообщалось, что cp184-mCherry демонстрирует 18% нативной флуоресценции mCherry, когда белки экспрессируются в E.coli, и 37% флуоресценции при сравнении изолированных белков [18]. Эволюция cp193-mCherry в cp193g7 (соответствующий cp-mCherry 197-196 ) путем случайного мутагенеза улучшила яркость этого варианта, достигнув 61% яркости mCherry на белковой основе [19].
Белковая яркость 6% по сравнению с mCherry (таблица 2). Разница между этими измерениями указывает на субоптимальную укладку белка, как обсуждается ниже. Ранее сообщалось, что cp184-mCherry демонстрирует 18% нативной флуоресценции mCherry, когда белки экспрессируются в E.coli, и 37% флуоресценции при сравнении изолированных белков [18]. Эволюция cp193-mCherry в cp193g7 (соответствующий cp-mCherry 197-196 ) путем случайного мутагенеза улучшила яркость этого варианта, достигнув 61% яркости mCherry на белковой основе [19].
10.1371/journal.pone.0020505.t001Таблица 1
| Вариант * | Яркость колонии через 24 ч при 37°C (% mCherry) | Яркость колонии через 72 ч при 2–8°C (% mCherry) |
| мЧерри | 100 | 100 |
| PRSET-A | 0 | 0 |
| cp99-98 | 0 | 0 |
| cp107-106 | 0 | 0 |
| cp119-118 | 0 | 0 |
| cp143-142 | 0 | 0 |
| cp149-148b | 0 | 0 |
| cp153-152 | 0 | 0 |
| Район петли 7–8 и фланговые участки | ||
| cp156-155 | 0 | 0 |
| cp157-156 | 0 | 0. 7 7 |
| cp158-157a | 0 | 2,8 |
| cp159-158 | 0,1 | 17,5 |
| cp160-159 | 0,1 | 12,3 |
| cp161-160 | 0 | 0,3 |
| cp162-161 | 0 | 0 |
| Район петли 8–9 и фланговые участки | ||
| cp171-170 | 0 | 0 |
| cp172-171 | 0 | 0. 4 4 |
| cp173-172 | 0 | 1,1 |
| cp174-173 | 0,1 | 6,4 |
| cp175-174a | 0,3 | 19,8 |
| cp176-175 | 0 | 3,9 |
| cp177-176 | 0 | 4. 3 3 |
| cp178-177 | 0 | 1,0 |
| cp179-178 | 0 | 0 |
| Район петли 9–10 и фланговые участки | ||
| cp188-187 | 0 | 0 |
| cp189-188 | 0 | 0. 7 7 |
| cp190-189 | 0,4 | 36,3 |
| cp191-190 | 0,3 | 19,8 |
| cp192-191 | 0 | 11,0 |
| cp193-192a | 0,2 | 17,2 |
| cp194-193 | 1. 0 0 |
39 |
| cp195-194 | 0 | 3,3 |
| cp196-195 | 0,2 | 3,1 |
| cp197-196 | 0,3 | 3,8 |
| cp198-197 | 0,1 | 3.8 |
| cp199-198 | 0 | 0 |
| Опубликовано cp-mCherryc | ||
| cp184 | <0,5 | 18 |
| cp193 | <0,5 | 1 |
| cp193g7 | 69 | 100 |
*
варианта mCherry с циклической пермутацией были пронумерованы по первичной аминокислотной последовательности mCherry, как показано на рис. 1С. Варианты помечены новыми амино- и карбокси-концами (например, cp159-158 имеет новый N-конец от нативных карбоксиаминокислот с 159 по 236 и новый С-конец от амино-остатков с 1 по 158).
1С. Варианты помечены новыми амино- и карбокси-концами (например, cp159-158 имеет новый N-конец от нативных карбоксиаминокислот с 159 по 236 и новый С-конец от амино-остатков с 1 по 158).
а
флуоресцентных варианта исходного скрининга в каждом регионе.
б
Соответствует cp-EGFP в GCaMP2.
с
Пронумерованы аминокислотной последовательностью DsRed, как показано на рис. 1C[18], [19].cp184-mCherry соответствует cp191-190 с тремя дублированными остатками на N-конце (188-190). cp193-mCherry и cp193g7 (с 6 мутированными аминокислотами) соответствуют cp197-196 с тремя дублированными остатками (197-199) на С-конце.
10.1371/journal.pone.0020505.t002Таблица 2
| Белок | Коэффициент экстинкцииb (M -1 см -1 ) | Квантовый выходc | Относительная яркостьd (% mCherry) | Яркость протеина (% mCherry) |
| мЧерри | 91 000f | 0. 22 22 |
100 | 100 |
| cp159-158 | 91 000 | 0,21 | 95 | 37,4 |
| cp160-159 | 89 000 | 0,21 | 93 | 22,0 |
| cp175-174 | 91 000 | 0. 20 20 |
91 | 41,5 |
| cp190-189 | 88 000 | 0,21 | 92 | 59,1 |
| cp191-190 | 93 000 | 0,20 | 93 | 53,3 |
| cp193-192 | 93 000 | 0. 20 20 |
93 | 51,7 |
| cp194-193 | 90 000 | 0,20 | 90 | 60,6 |
| cp184 | 26 600 г | 0,22 | НДи | 37,0 |
| cp193g7 | 42 000 г | 0. 23 23 |
НДи | 60,6 |
а
Все варианты с круговой пермутацией из нативной последовательности mCherry, показанные в этой таблице, за исключением cp184 (не определено), имеют такие же максимумы возбуждения (587 нм) и испускания (610 нм), что и mCherry. Максимумы возбуждения и испускания cp193g7 с 6 аминокислотными мутациями составляют 580 нм и 602 нм соответственно [19].
б
Коэффициенты экстинкции измерены методом щелочно-денатурированных хромофоров.
с
Квантовый выход измеряли с использованием mCherry в качестве стандарта сравнения.
д
Относительную яркость хромофора (коэффициент экстинкции × квантовый выход) сравнивали с mCherry (
е
Флуоресценция cp-mCherry по сравнению с mCherry с фиксированной концентрацией белка (анализ BCA).
ф
Наши данные; опубликованные данные 72 000 [7], 78 000 [9].
г
Опубликованные значения [18], [19], основанные на количественном определении белка (поглощение при 280 нм).
я
Не определено.
Циклическая перестановка mKate
Мы провели аналогичный циклический анализ перестановки мономерного дальнекрасного белка mKate, который происходит от актинии Entacmaea quadricolor и имеет структурное сходство с mCherry. Основываясь на сходстве структур и последовательностей, мы сначала протестировали три варианта cp-mKate, которые соответствовали наиболее ярким вариантам cp-mCherry в каждой области пермиссивных сайтов.Флуоресценция была обнаружена в колониях, трансформированных cp-mKate 151-150 , cp-mKate 167-166 и cp-mKate 189-188 . Как и в случае оценки cp-mCherry, мы также проверили последовательности, фланкирующие три идентифицированных круговых сайта перестановки в mKate, на наличие дополнительных толерантных сайтов. В отличие от медленного развития флуоресценции в колониях cp-mCherry, большинство флуоресцентных вариантов cp-mKate демонстрировали заметную флуоресценцию после 24-часовой инкубации при 37°C (табл. 3).cp-mKate 189-188 — самый яркий вариант круговой пермутации у E.coli. Однако анализ очищенных белков в 20 мМ HEPES (pH 7,4) показал, что яркость cp-mKate 149-148 , mKate 151-150 , cp-mKate 167-166 и cp-mKate 168 -167 был довольно высоким, превышая 80% флуоресценции нативного mKate, что указывает на высокоэффективные флуоресцентные конфигурации. Пик поглощения при 442 нм (рис. 3В), который испускает зеленую флуоресценцию при 532 нм, выявил небольшое усиление зеленой флуоресценции в cp-mKates.Соотношение поглощения при 588 нм и 442 нм (табл. 4) указывало на относительно высокий процент зеленых компонентов в cp-mKate 154-153 , cp-mKate 168-167 , cp-mKate 187-186 , и cp-mKate 189-188 , тогда как cp-mKate 149-148 демонстрирует спектр флуоресценции, аналогичный нативному mKate.
В отличие от медленного развития флуоресценции в колониях cp-mCherry, большинство флуоресцентных вариантов cp-mKate демонстрировали заметную флуоресценцию после 24-часовой инкубации при 37°C (табл. 3).cp-mKate 189-188 — самый яркий вариант круговой пермутации у E.coli. Однако анализ очищенных белков в 20 мМ HEPES (pH 7,4) показал, что яркость cp-mKate 149-148 , mKate 151-150 , cp-mKate 167-166 и cp-mKate 168 -167 был довольно высоким, превышая 80% флуоресценции нативного mKate, что указывает на высокоэффективные флуоресцентные конфигурации. Пик поглощения при 442 нм (рис. 3В), который испускает зеленую флуоресценцию при 532 нм, выявил небольшое усиление зеленой флуоресценции в cp-mKates.Соотношение поглощения при 588 нм и 442 нм (табл. 4) указывало на относительно высокий процент зеленых компонентов в cp-mKate 154-153 , cp-mKate 168-167 , cp-mKate 187-186 , и cp-mKate 189-188 , тогда как cp-mKate 149-148 демонстрирует спектр флуоресценции, аналогичный нативному mKate. Квантовый выход при 588 нм для cp-mKate лишь немного ниже, чем для mKate, тогда как соотношение A 588 / A 280 указывает на то, что ряд молекул cp-mKate (cp-mKate 154-153 , cp-mKate 187-186 , cp-mKate 189-188 ) имеют значительно более низкое поглощение при 588 нм при соответствующей концентрации белка (таблица 4).
Квантовый выход при 588 нм для cp-mKate лишь немного ниже, чем для mKate, тогда как соотношение A 588 / A 280 указывает на то, что ряд молекул cp-mKate (cp-mKate 154-153 , cp-mKate 187-186 , cp-mKate 189-188 ) имеют значительно более низкое поглощение при 588 нм при соответствующей концентрации белка (таблица 4).
10.1371/journal.pone.0020505.g003Рисунок 3
спектра нормализовали по поглощению при 280 нм для каждого белка. А. Спектры поглощения вариантов mCherry и cp-mCherry. B. Спектры поглощения вариантов mKate и cp-mKate.
10.1371/journal.pone.0020505.t003Таблица 3
| Вариант * | Яркость колонии через 24 ч при 37°C (% mKate) | Яркость колонии через 72 ч при 2–8°C (% mKate) |
| мКейт | 100 | 100 |
| PRSET-A | 0 | 0 |
| cp139-138a | 0 | 0 |
| цп140-139а | 0 | 0 |
| ср141-140а | 0 | 0 |
| cp144-143a | 0 | 0 |
| Район петли 7-8 и фланговые участки | ||
| cp148-147 | 0 | 0 |
| cp149-148 | 1. 1 1 |
32,3 |
| cp150-149 | 0,2 | 11,7 |
| cp151-150b | 7,5 | 28,4 |
| cp152-151 | 5,1 | 21,4 |
| cp153-152 | 2,9 | 20.8 |
| cp154-153 | 10,9 | 19,4 |
| cp155-154 | 0 | 0 |
| cp156-155 | 0 | 0 |
| Район петли 8–9 и фланговые участки | ||
| cp164-163 | 0 | 0 |
| cp165-164 | 1. 9 9 |
13,2 |
| cp166-165 | 10,2 | 24,7 |
| cp167-166c | 6,6 | 21,6 |
| cp168-167 | 3,6 | 22,2 |
| cp169-168 | 1,6 | 16.3 |
| cp170-169 | 0 | 0 |
| Район петли 9–10 и фланговые участки | ||
| cp180-179 | 0 | 0 |
| cp181-180 | 0 | 0 |
| cp182-181 | 11. 5 5 |
12,6 |
| cp183-182 | 23,7 | 30,0 |
| cp184-183 | 18,7 | 28,4 |
| cp185-184 | 9,7 | 17,5 |
| cp186-185 | 21,2 | 27.7 |
| cp187-186 | 26,7 | 30,8 |
| cp188-187 | 9,9 | 19,7 |
| cp189-188d | 33,9 | 55,2 |
| cp190-189 | 12,1 | 18,3 |
| cp191-190 | 8. 3 3 |
15 |
| cp192-191 | 0,4 | 6 |
| cp193-192 | 0 | 0 |
| cp194-193 | 0 | 0 |
*
Варианты с круговой перестановкой помечены новыми амино- и карбокси-концами.
а
Выравнивание по cp-EGFP в GCaMP2.
б
Выравнивает cp-mCherry 159-158 .
с
Выравнивает cp-mCherry 175-174 .
д
Выравнивает cp-mCherry 194-193 .
10.1371/journal.pone.0020505.t004Таблица 4
| Белок | Поглощениеb (A 588 /A 442 ) | Квантовый выходc (588 нм) | Поглощенный (A 588 /A 280 ) | Яркость протеина (% mKate) |
| мКейт | 4. 13 13 |
0,33 | 0,74 | 100 |
| cp149-148 | 4,79 | 0,33 | 0,68 | 90,1 |
| cp151-150 | 3,25 | 0,31 | 0,61 | 83,6 |
| cp154-153 | 0. 81 81 |
0,28 | 0,19 | 21,2 |
| cp166-165 | 2,88 | 0,28 | 0,58 | 70,5 |
| cp167-166 | 2,73 | 0,32 | 0,61 | 81,6 |
| cp168-167 | 1. 8 8 |
0,30 | 0,65 | 84,0 |
| cp187-186 | 1,45 | 0,30 | 0,25 | 30,4 |
| cp189-188 | 1,47 | 0,29 | 0,27 | 36,3 |
а
Красная флуоресценция всех вариантов mKate с круговой пермутацией, показанных в этой таблице, с N-концевым лидерным пептидом из вектора pRSET, имеет те же максимумы возбуждения (588 нм) и испускания (620 нм), что и mKate; тогда как опубликованный максимум эмиссии mKate составляет 635 нм [9] и 625 нм [24].
б
Максимумы поглощения красного и зеленого хромофоров составляют 588 нм и 442 нм соответственно.
с
Квантовый выход красной флуоресценции при возбуждении 588 нм измеряли с использованием mKate в качестве стандарта сравнения.
д
Соотношение поглощения красного хромофора при 588 нм и поглощения белка при 280 нм.
е
Флуоресценция cp-mKate относительно mKate с фиксированной концентрацией белка (анализ BCA).
Мы также провели скрининг вариантов cp-mKate, которые соответствуют сайту в cp-EGFP, используемому для разработки семейства сенсоров кальция GCaMP [2], [3], на основе структурного и последовательного выравнивания mKate и EGFP. Колонии, трансформированные с помощью cp-mKate 139-138 , cp-mKate 140-139 , cp-mKate 141-140 , cp-mKate 144-143 , не проявляли красной флуоресценции, даже когда инкубировались сут при 2–8°С (табл. 3).
3).
Усечения cp-mKate
Чтобы изучить возможность влияния N- или C-концевых остатков на флуоресценцию отдельных круговых перестановок, мы затем определили чувствительность флуоресценции к делециям на амино- и карбокси-концах флуоресцентных пермутантов cp-mKate. .Был сконструирован ряд вариантов с укороченными амино- или карбокси-концами. В этом отношении индивидуальную конструкцию можно рассматривать как амино- или карбоксиделецию; таким образом, cp-mKate 154-150 , например, представляет собой амино-укорочение cp-mKate 151-150 или карбокси-усечение cp-mKate 154-153 . Как показано в таблице 3, для области петли 9–10 cp-mKates флуоресцентные варианты содержат N-концевой остаток от 182 до 192 и С-концевой остаток от 181 до 191 соответственно).Большинство укороченных вариантов cp-mKate 183-182 и cp-mKate 182-181 заметно снижали флуоресценцию в E.coli, за исключением cp-mKate 185-182 , эффективность которого была аналогична cp-mKate 183-182 (табл. 5). N- и C-концевые границы для усечения, при которых сохранялась флуоресценция, составляли 192 и 181 соответственно, что указывает на то, что усечения в области круговой перестановки допустимы. Аналогичные эксперименты были проведены в области вариантов cp-mKate петли 7–8 и петли 8–9, что указывает на то, что амино- и карбоксильные границы для укорочения представляют собой остатки 154 и 148 для области петли 7–8 и 169 и 164 для петли 8. –9 регион (данные не показаны).
5). N- и C-концевые границы для усечения, при которых сохранялась флуоресценция, составляли 192 и 181 соответственно, что указывает на то, что усечения в области круговой перестановки допустимы. Аналогичные эксперименты были проведены в области вариантов cp-mKate петли 7–8 и петли 8–9, что указывает на то, что амино- и карбоксильные границы для укорочения представляют собой остатки 154 и 148 для области петли 7–8 и 169 и 164 для петли 8. –9 регион (данные не показаны).
10.1371/journal.pone.0020505.t005Таблица 5
| Вариант | Яркость колонии через 24 ч при 37°C (% mKate) | Яркость колонии через 72 ч при 2–8°C (% mKate) |
| cp183-182* | 21,4 | 24,6 |
| cp185-182 | 23. 1 1 |
24,2 |
| cp188-182 | 15,1 | 13,3 |
| cp191-182 | 7,9 | 11,5 |
| cp193-182 | 0 | 0 |
| cp182-181* | 16,9 | 16.9 |
| cp186-181 | 2,6 | 2,2 |
| cp191-181 | 1,8 | 3,1 |
| cp192-181 | 0,2 | 1 |
| cp193-181 | 0 | 0 |
| cp181-180* | 0 | 0 |
| PRSET-A | 0 | 0 |
| мКейт | 100 | 100 |
*Оригинальный симметричный cp-mKate.
Кристаллическая структура cp-mKate
Рентгеновская кристаллография на cp-mKate 154-153 и cp-mKate 168-167 позволила нам определить структуру двух пермутантов с разрешением 3,0 Å и 1,7 Å соответственно (таблица). 6 и рис. 4). Разрешенная структура очень похожа на ту, о которой сообщалось ранее для mKate дикого типа [13] (среднее квадратичное отклонение маршрута 0,31 Å и 0,23 Å соответственно), обнаруживая эллиптическую β-бочку, которая правильно свернута в cp-mKate 168- 167 и неотличимы по третичной структуре от структуры mKate дикого типа (рис.4А). Линкер, который соединяет два переориентированных компонента молекулы с круговой перестановкой, появляется в петлевой структуре и располагается на одном конце β-ствола (Fig. 4A). Незначительные конформационные изменения в cp-mKate были ограничены петлевыми структурами на обоих концах β-ствола и могут объяснить зарегистрированный красно-зеленый сдвиг. Однако хромофор полностью находится в цис-конформации с водородными связями с Trp 90, Arg92 и Ser143 (Fig. 4B).
4B).
10.1371/журнал.поне.0020505.g004Рисунок 4
A. Мультяшная презентация mKate (pH 7,0), cp-mKate 154-153 и cp-mKate 168-167 . Фигуры нарисованы с помощью PyMol (DeLano Scientific). B. Карта электронной плотности 2mFo-Fc вблизи области хромофора. Карта оконтурена на 1.0σ.
10.1371/journal.pone.0020505.t006Таблица 6
| cp-mKate 154-153 | cp-mKate 168-167 | |
| Сбор данных | ||
| Источник рентгеновского излучения | ШАХМАТЫ A1 | ШАХМАТЫ A1 |
| Длина волны (Å) | 0,978 | 0. 978 978 |
| Космическая группа | P2 1 2 1 2 1 | С2 |
| Параметры элементарной ячейки | ||
| а, б, в (Å) | 71,46, 71,44, 367,73 | 109,357, 93,280,194,370 |
| α, β, γ (°) | 90,0, 90.0, 90,0 | 90,0, 96,52, 90,0 |
| Разрешение (Å) | 38,98-3,00 (3,11-3,00) | 38,31-1,75 (1,81-1,75) |
| Количество отражений | ||
| Итого | 330773 (32752) | 180131 (15380) |
| Уникальный | 37589 (3680) | 45032 (4660) |
| Полнота (%) | 99. 9 (100,0) 9 (100,0) |
92,5 (79,6) |
| Резервирование | 8,8 (8,9) | 4,0 (3,3) |
| I/σ(I) | 15,3 (2,0) | 39,4 (7,26) |
| R изм. (%) | 11,9 (47,7) | 5,1 (14,5) |
| Уточнение | ||
| Р работа /Р бесплатно (%) | 21. 7/27.8 7/27.8 |
18,0/21,1 |
| среднеквадратичное значение отклонения | ||
| Длина связи (Å) | 0,009 | 0,007 |
| Валентные углы (°) | 1,590 | 1,164 |
| Количество атомов | ||
| Белок | 14974 | 16094 |
| Вода | 2 | 1453 |
Обсуждение
Генерация большого количества круговых перестановок с помощью тандемной матричной ПЦР и флуоресцентного скрининга бактериальных колоний является высокоэффективным подходом к изучению потенциальных вариантов перестановок флуоресцентных белков. Мы систематически исследовали mCherry и mKate, создавая круговые перестановки в областях петель, в которых можно было бы ожидать наибольшей гибкости. При попытке разработать высокофлуоресцентный красный белок с круговой пермутацией стратегии дизайна, направленные на структурную имитацию cp-EGFP, которая является основой для успешных сенсоров GCaMP Ca 2+ [3], [20], [21], оказались неэффективными. быть успешным подходом, поскольку аналогичные варианты cp как mCherry, так и mKate не показали значительной флуоресценции.Структурный анализ выявил три высоко гомологичных участка петель у mCherry и mKate, и эти области вместе с фланкирующими участками были систематически исследованы. Каждая область допускала циркулярную перестановку, но флуоресценция заметно различалась, при этом наиболее эффективные участки перестановки имели тенденцию возникать в пределах центральной области петли.
Мы систематически исследовали mCherry и mKate, создавая круговые перестановки в областях петель, в которых можно было бы ожидать наибольшей гибкости. При попытке разработать высокофлуоресцентный красный белок с круговой пермутацией стратегии дизайна, направленные на структурную имитацию cp-EGFP, которая является основой для успешных сенсоров GCaMP Ca 2+ [3], [20], [21], оказались неэффективными. быть успешным подходом, поскольку аналогичные варианты cp как mCherry, так и mKate не показали значительной флуоресценции.Структурный анализ выявил три высоко гомологичных участка петель у mCherry и mKate, и эти области вместе с фланкирующими участками были систематически исследованы. Каждая область допускала циркулярную перестановку, но флуоресценция заметно различалась, при этом наиболее эффективные участки перестановки имели тенденцию возникать в пределах центральной области петли.
Как показано в Таблице 1 и Таблице 2, самый яркий вариант mCherry (cp-mCherry 194-193 ) сохранил 60,6% яркости нативного mCherry на белковой основе, но также наблюдалась сильная флуоресценция для круговых перестановок cp-Cherry. 159-158 , CP-Cherry 160-159 , CP-MCRCER 175-174 , CP-MCHERR 190-189 , CP-MCRERY 191-190 и CP-MCHERR 193-192 .Все варианты cp-mCherry показали такие же спектры поглощения, как и нативный mCherry (рис. 3А). Более того, красный хромофор в вариантах cp-mCherry и mCherry функционально подобен, имея почти одинаковые значения коэффициента экстинкции и квантового выхода. Таким образом, заметная разница в относительной яркости между вариантами cp-mCherry и нативной mCherry при оценке на основе хромофора и эквивалентной концентрации белка указывает на то, что снижение яркости белков cp-mCherry связано с неполным сворачиванием значительной части белка. .Эта интерпретация также подтверждается тем фактом, что 6 аминокислотных мутаций в cp193g7, которые, как было показано, улучшают эффективность фолдинга белка, приводили к гораздо более высокой флуоресценции, чем наблюдаемая в cp193-mCherry [19], сходной с эффектом яркости в случае cp193-mCherry [19].
159-158 , CP-Cherry 160-159 , CP-MCRCER 175-174 , CP-MCHERR 190-189 , CP-MCRERY 191-190 и CP-MCHERR 193-192 .Все варианты cp-mCherry показали такие же спектры поглощения, как и нативный mCherry (рис. 3А). Более того, красный хромофор в вариантах cp-mCherry и mCherry функционально подобен, имея почти одинаковые значения коэффициента экстинкции и квантового выхода. Таким образом, заметная разница в относительной яркости между вариантами cp-mCherry и нативной mCherry при оценке на основе хромофора и эквивалентной концентрации белка указывает на то, что снижение яркости белков cp-mCherry связано с неполным сворачиванием значительной части белка. .Эта интерпретация также подтверждается тем фактом, что 6 аминокислотных мутаций в cp193g7, которые, как было показано, улучшают эффективность фолдинга белка, приводили к гораздо более высокой флуоресценции, чем наблюдаемая в cp193-mCherry [19], сходной с эффектом яркости в случае cp193-mCherry [19]. зеленый флуоресцентный белок superfolder [22].
зеленый флуоресцентный белок superfolder [22].
В целом флуоресценция пермутантов cp-mCherry из нативного mCherry развивалась медленнее и достигала гораздо большей яркости при дальнейшей инкубации бактерий при более низкой температуре.Напротив, варианты cp-mKate проявляли значительную флуоресценцию в бактериях через 24 часа при 37°C (таблица 3). Самым ярким вариантом в бактериальном анализе был cp-mKate 189-188 , при этом колонии достигали 55,2% яркости нативного mKate через 72 часа инкубации. Однако яркость бактериальных колоний не коррелировала строго с флуоресценцией белка, и белки, выделенные только из умеренно флуоресцентных колоний, были одними из самых ярких идентифицированных белков. Таким образом, хотя cp-mKate 189-188 продемонстрировал самую яркую флуоресценцию колонии, очищенный белок показал 36.3% нативной флуоресценции mKate, тогда как относительная флуоресценция cp-mKate 149-148 , cp-mKate 151-150 , cp-mKate 167-166 и cp-mKate 168-167 % при рН 7,4. Более высокая флуоресценция этих белков может быть связана с цитозольными факторами, такими как pH, что приводит к более высокому проценту правильно свернутого белка и зрелого красного хромофора в очищенных белках.
Более высокая флуоресценция этих белков может быть связана с цитозольными факторами, такими как pH, что приводит к более высокому проценту правильно свернутого белка и зрелого красного хромофора в очищенных белках.
Ранее сообщалось об отдельных сайтах модификации mKate, допускающих циклическую перестановку или расщепление.В пробе напряжения использовался cp-mKate(180) [23], который содержал 3 дублированные аминокислоты на N-конце и в остальном соответствовал умеренно яркому cp-mKate 183-182 . Сообщаемый сайт функционального расщепления для mKate (151) для системы BIFC [24] принадлежит сильно флуоресцентной области петли 7-8 (таблица 3). Мы также подтвердили устойчивость выбранных сайтов к вставкам пептидов, которая оказалась надежной (данные не показаны).
Хотя mKate и его варианты широко используются в качестве красных флуоресцентных белков, эти белки проявляют зеленую флуоресценцию в различной степени [25].Мы обнаружили, что круговая перестановка mKate усиливала зеленую флуоресценцию в нескольких конструкциях, увеличивая поглощение при 442 нм. Поскольку любая зеленая флуоресценция белков mKate будет вносить вклад в общую флуоресценцию после щелочной денатурации, точное измерение коэффициента экстинкции красного хромофора в cp-mKate с помощью этого метода недопустимо. Как и в случае круговых пермутантов mCherry, квантовый выход красной флуоресценции лишь немного снижается в cp-mKates. В случае mCherry коэффициенты экстинкции белков с круговой пермутацией и белков дикого типа очень похожи, а уменьшение отношения поглощения A 587 /A 280 указывает на то, что вклад плохо свернутых белков является основным фактором в потеря яркости (таблица 2 и рис.3А). Однако для вариантов mKate с меньшей яркостью, чем у нативного белка, сходные соотношения A 588 /A 280 между этими формами отражают не только эффект неправильно свернутого белка, но и степень, в которой перестановка привела к красно-зеленому цвету. сдвиг. Интересно, что мы обнаружили очень яркие cp-mKate со значительным красно-зеленым сдвигом, такие как cp-mKate 168-167 , и очень яркие варианты без существенного сдвига, такие как cp-mKate 149-148 (табл.
Поскольку любая зеленая флуоресценция белков mKate будет вносить вклад в общую флуоресценцию после щелочной денатурации, точное измерение коэффициента экстинкции красного хромофора в cp-mKate с помощью этого метода недопустимо. Как и в случае круговых пермутантов mCherry, квантовый выход красной флуоресценции лишь немного снижается в cp-mKates. В случае mCherry коэффициенты экстинкции белков с круговой пермутацией и белков дикого типа очень похожи, а уменьшение отношения поглощения A 587 /A 280 указывает на то, что вклад плохо свернутых белков является основным фактором в потеря яркости (таблица 2 и рис.3А). Однако для вариантов mKate с меньшей яркостью, чем у нативного белка, сходные соотношения A 588 /A 280 между этими формами отражают не только эффект неправильно свернутого белка, но и степень, в которой перестановка привела к красно-зеленому цвету. сдвиг. Интересно, что мы обнаружили очень яркие cp-mKate со значительным красно-зеленым сдвигом, такие как cp-mKate 168-167 , и очень яркие варианты без существенного сдвига, такие как cp-mKate 149-148 (табл. 4 и Инжир.3Б). Общая яркость этих конструкций свидетельствует о минимальных эффектах неполной укладки при рН 7,4, тогда как потеря яркости в вариантах с аналогичными красно-зелеными сдвигами наблюдается, например, при сравнении cp-mKate 168–167 и cp-mKate 187. -186 , указывает на менее эффективную укладку и стабилизацию хромофора в последнем варианте (табл. 4 и рис. 3В). cp-mKate 149-148 был самым ярким белком, идентифицированным с более чем 90 процентами естественной яркости, но с 3 дополнительными вариантами (cp-mKate 151-150 , cp-mKate 167-166 и cp-mKate 168). -167 ) превышают 80% флуоресценции mKate, составляя новые пермутанты, доступные для реализации в качестве сенсорных или комплементарных пар.Кроме того, открытие ярких сенсоров с существенной зеленой флуоресценцией, таких как cp-mKate 168-167 или стабилизированный cp-mKate 154-153 , может быть полезным при разработке зелено-красных логометрических сенсоров.
4 и Инжир.3Б). Общая яркость этих конструкций свидетельствует о минимальных эффектах неполной укладки при рН 7,4, тогда как потеря яркости в вариантах с аналогичными красно-зелеными сдвигами наблюдается, например, при сравнении cp-mKate 168–167 и cp-mKate 187. -186 , указывает на менее эффективную укладку и стабилизацию хромофора в последнем варианте (табл. 4 и рис. 3В). cp-mKate 149-148 был самым ярким белком, идентифицированным с более чем 90 процентами естественной яркости, но с 3 дополнительными вариантами (cp-mKate 151-150 , cp-mKate 167-166 и cp-mKate 168). -167 ) превышают 80% флуоресценции mKate, составляя новые пермутанты, доступные для реализации в качестве сенсорных или комплементарных пар.Кроме того, открытие ярких сенсоров с существенной зеленой флуоресценцией, таких как cp-mKate 168-167 или стабилизированный cp-mKate 154-153 , может быть полезным при разработке зелено-красных логометрических сенсоров.
варианта усечения показали, что в каждой области cp-mKate имеется минимум нативных N-концевых и C-концевых фрагментов, необходимых для поддержания флуоресценции. Например, минимальные С-концевые и N-концевые фрагменты составляли 192–231 и 1–181 для cp-mKate области петли 9–10.Способность укорачивать варианты без существенной потери флуоресценции обеспечивает значительную гибкость связывания этих пермутантов с другими функциональными пептидами.
Наконец, кристаллизация и структурный анализ cp-mKate 154-153 и cp-mKate 168-167 выявили ожидаемую третичную структуру, о которой ранее сообщалось для mKate [16], с небольшими вариациями вокруг точки перестановки. Будущие исследования будут направлены на определение структурной основы для изменения флуоресценции в кольцевых пермутантах mKate.
Таким образом, мы сообщаем о нескольких сильно флуоресцентных вариантах с круговой пермутацией mCherry и mKate. Эти варианты сгруппированы в 3 области и представляют собой самые ярко-красные циркулярно пермутированные белки с нативными белковыми последовательностями, о которых сообщалось на сегодняшний день. Сообщаемые белки mKate с яркой круговой пермутацией и дополнительно стабилизированные варианты mCherry должны предоставить дополнительных кандидатов для конструирования красных сенсоров и инструментов комплементации.
Сообщаемые белки mKate с яркой круговой пермутацией и дополнительно стабилизированные варианты mCherry должны предоставить дополнительных кандидатов для конструирования красных сенсоров и инструментов комплементации.
Номера доступа
Атомные координаты и структурные факторы были депонированы в Банк данных белков RCSB под идентификационными кодами 3rwt и 3rwa.
Экспериментальная и вычислительная электрофизиология с высоким разрешением выявляет слабое связывание β-лактамов в поринах PorB
Delcour, A. H. Поглощение растворенных веществ через общие порины. Фронт. Бионауч. 8 , 1055–1071 (2003).
Артикул
Google Scholar
Delcour, A. H. Проницаемость наружной мембраны и устойчивость к антибиотикам. Биохим. Биофиз. Acta 1794 , 808–16 (2009).
КАС
пабмед
Статья
Google Scholar
Никайдо, Х. Молекулярные основы проницаемости внешней мембраны бактерий. Микробиолог. Мол. биол. 67 , 593–656 (2003).
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
James, C.E. et al . Как β-лактамные антибиотики проникают в бактерии: диалог с поринами. PLoS One 4 , e5453 (2009 г.).
ОБЪЯВЛЕНИЕ
пабмед
ПабМед Центральный
Статья
Google Scholar
Pages, J.-M., James, C.E. & Winterhalter, M. Порин и проникающий антибиотик: селективный диффузионный барьер для грамотрицательных бактерий. Нац. Преподобный Микробиолог. 6 , 893–903 (2008).
ПабМед
Статья
Google Scholar
Simonet, V., Malléa, M. & Pagès, J.M. Замены в области глазка нарушают диффузию цефепима через канал OmpF Escherichia coli. Антимикроб. Агенты Чемотер. 44 , 311–315 (2000).
Антимикроб. Агенты Чемотер. 44 , 311–315 (2000).
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Бредин, Дж. и др. . Изменение пористости OmpF Escherichia coli, вызванное мутацией ключевых остатков в области анти-петли 3. Биохим. J. 363 , 521–8 (2002).
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Олески, М., Хоббс, М. и Николас, Р. А. Идентификация и анализ аминокислотных мутаций в порине IB, которые опосредуют промежуточную устойчивость к пенициллину и тетрациклину у Neisseria gonorrhoeae. Антимикроб. Агенты Чемотер. 46 , 2811–2820 (2002).
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Баджадж, Х. и др. . Молекулярные основы фильтрации карбапенемов поринами из резистентных к бета-лактаму клинических штаммов кишечной палочки. J. Biol. хим. 291 , 2837–2847 (2016).
J. Biol. хим. 291 , 2837–2847 (2016).
КАС
пабмед
Статья
Google Scholar
Piddock, L. J. V. Понимание основ устойчивости к антибиотикам: платформа для открытия лекарств. Микробиология 2366–2373, https://doi.org/10.1099/мик.0.082412-0 (2014).
Льюис, К. Платформы для открытия антибиотиков. Нац. Преподобный Друг Дисков. 12 , 371–387 (2013).
КАС
пабмед
Статья
Google Scholar
Карпентер, Т. С., Паркин, Дж. и Халид, С. Свободная энергия проникновения малых растворенных веществ через внешнюю мембрану кишечной палочки имеет отчетливо асимметричный профиль. J. Phys. хим. лат. 7 , 3446–3451 (2016).
КАС
пабмед
Статья
Google Scholar
Хсу, П.-К., Джеффрис, Д. и Халид, С. Молекулярно-динамическое моделирование прогнозирует пути проникновения первичных фуллеренов через бактериальные мембраны. J. Phys. хим. B 120 , 11170–11179 (2016).
J. Phys. хим. B 120 , 11170–11179 (2016).
КАС
пабмед
Статья
Google Scholar
ВОЗ. ВОЗ публикует список бактерий, для которых срочно необходимы новые антибиотики (2017 г.).
Пендлтон, Дж. Н., Горман, С. П. и Гилмор, Б. Ф. Клиническая значимость патогенов ESKAPE. Эксперт Преподобный Анти. Заразить. тер. 11 , 297–308 (2013).
КАС
пабмед
Статья
Google Scholar
Батлер, М. С., Бласкович, М. А. и Купер, М. А. Антибиотики в клинической разработке в конце 2015 г. Дж. Антибиот. (Токио). 70 , 3–24 (2017).
КАС
пабмед
Статья
Google Scholar
Theuretzbacher, U. Инновационные антибиотики для будущих потребностей общественного здравоохранения. клин. микробиол. инфекц. 23 , 713–717 (2017).
КАС
Статья
Google Scholar
Сильвер, Л. Л. Проблемы открытия антибактериальных средств. клин. микробиол. 24 , 71–109 (2011).
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Всемирная организация здравоохранения. Антибактериальные препараты в клинической разработке: анализ хода клинической разработки антибактериальных препаратов, включая туберкулез. WHO/EMP/IAU/2017.12 48 (2017).
Несторович Э.М., Данелон К., Винтерхальтер М.и Безруков С. М. Предназначен для проникновения: взаимодействие одиночных молекул антибиотика с бактериальными порами с временным разрешением. Проц. Натл. акад. науч. США 99 , 9789–9794 (2002).
ОБЪЯВЛЕНИЕ
КАС
пабмед
Статья
Google Scholar
Райт, Г. Д. и др. . Молекулярные механизмы устойчивости к антибиотикам. Хим. коммун. 47 , 4055–4061 (2011).
. Молекулярные механизмы устойчивости к антибиотикам. Хим. коммун. 47 , 4055–4061 (2011).
КАС
Статья
Google Scholar
Рихтер, М.Ф. и др. . Правила прогнозирования накопления соединений позволяют получить антибиотик широкого спектра действия. Природа 545 , 299–304 (2017).
ОБЪЯВЛЕНИЕ
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Пейн, Ф., Текуапетла-Гомес, И., Шютте, О. М., Стейнем, К. и Мунк, А. Полностью автоматическая многоразрешающая идеализация для отфильтрованных записей ионных каналов: обнаружение мерцающих событий. Транзакция IEEE. нанобиоск. 17 , 300–320 (2018).
Танабе, М., Нимигеан, К.М. и Айверсон, Т.М. Структурные основы транспорта растворенных веществ, регуляция нуклеотидов и иммунологическое распознавание Neisseria meningitidis PorB. Проц. Натл. акад. науч. США 107 , 6811–6 (2010).
Проц. Натл. акад. науч. США 107 , 6811–6 (2010).
ОБЪЯВЛЕНИЕ
КАС
пабмед
Статья
Google Scholar
Каттнер, К. и др. . Кристаллографический анализ внеклеточных петель PorB Neisseria meningitidis, потенциально участвующих в распознавании TLR2. Дж. Структура. биол. 185 , 440–447 (2014).
КАС
пабмед
Статья
Google Scholar
Массари, П. и др. . Менингококковый порин PorB связывается с TLR2 и требует TLR1 для передачи сигналов. Дж. Иммунол. 176 , 2373–2380 (2006).
КАС
пабмед
Статья
Google Scholar
Пик, И. Р., Дженнингс, К. Д., Джен, Ф. Е. К. и Дженнингс, М. П. Роль экспрессии neisseria meningitidis pora и porB в чувствительности к противомикробным препаратам. Антимикроб. Агенты Чемотер. 58 , 614–615 (2014).
58 , 614–615 (2014).
ПабМед
ПабМед Центральный
Статья
Google Scholar
Оппенгейм, Б.A. Антибиотикорезистентность Neisseria meningitidis. клин. Заразить. Дис. 24 , 98–101 (1997).
Артикул
Google Scholar
Kattner, C., Zaucha, J., Jaenecke, F., Zachariae, U. & Tanabe, M. Идентификация пути транспорта катионов в Neisseria meningitidis PorB. Структура белков. Функц. Биоинформа. 81 , 830–840 (2013).
КАС
Статья
Google Scholar
Zachariae, U., Helms, V. & Engelhardt, H. Многоступенчатый механизм переноса хлоридов в сильно анионселективном пориновом канале. Биофиз. J. 85 , 954–962 (2003).
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar

Ziervogel, B.K. & Roux, B. Связывание антибиотиков с OmpF-порином. Структура 21 , 76–87 (2013).
КАС
пабмед
Статья
Google Scholar
Махендран, К.Р. и др. . Молекулярная основа транслокации энрофлоксацина через OmpF, канал наружной мембраны кишечной палочки — При связывании транслокация не подразумевается. J. Phys. хим. B 114 , 5170–5179 (2010).
КАС
пабмед
Статья
Google Scholar
Данелон С., Несторович Э. М., Винтерхальтер М., Чеккарелли М. и Безруков С. М. Взаимодействие цвиттерионных пенициллинов с каналом OmpF облегчает их транслокацию. Биофиз. J. 90 , 1617–1627 (2006).
ОБЪЯВЛЕНИЕ
КАС
пабмед
Статья
Google Scholar
Бхамидимарри, С.П., Праджапати, Дж.Д., Ван Ден Берг, Б., Винтерхальтер, М. и Кляйнекатхофер, У. Роль электроосмоса в проникновении нейтральных молекул: CymA и циклодекстрин в качестве примера. Биофиз. J. 110 , 600–611 (2016).
и Кляйнекатхофер, У. Роль электроосмоса в проникновении нейтральных молекул: CymA и циклодекстрин в качестве примера. Биофиз. J. 110 , 600–611 (2016).
ОБЪЯВЛЕНИЕ
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Гаи, И. и др. . Общий метод определения потока заряженных молекул через нанопоры применительно к ингибиторам β-лактамаз и OmpF. J. Phys. хим. лат. 8 , 1295–1301 (2017).
КАС
пабмед
Статья
Google Scholar
Олески, М., Чжао, С., Розенберг, Р. Л. и Николас, Р. А. Порин-опосредованная устойчивость к антибиотикам у Neisseria gonorrhoeae: проникновение ионов, растворенных веществ и антибиотиков через белки PIB с мутациями penB. J. Бактериол. 188 , 2300–2308 (2006 г.).
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Song, J., Minetti, C.A.S.A., Blake, M.S. & Colombini, M. Успешное восстановление нормальных электрофизиологических свойств порина PorB (класс 3) из neisseria meningitidis после экспрессии в Escherichia coli и ренатурации. Биохим. Биофиз. Акта — биомембрана. 1370 , 289–298 (1998).
КАС
Статья
Google Scholar
Сугинта В., Винтерхальтер М. и Смит М.Ф. Коррелированное улавливание молекул сахара тримерным белковым каналом хитопорином. Биохим. Биофиз. Акта — биомембрана. 1858 , 3032–3040 (2016).
КАС
Статья
Google Scholar
Бодренко И., Баджай Х., Руджероне П., Винтерхальтер М.и Чеккарелли, М. Анализ блокировки быстрых каналов: выявление связывания субстрата в микросекундном диапазоне. Аналитик 140 , 4820–4827 (2015).
ОБЪЯВЛЕНИЕ
КАС
пабмед
Статья
Google Scholar
Бодренко И.В., Ван Дж., Салис С., Винтерхальтер М. и Чеккарелли М. Обнаружение проникновения одиночной молекулы в нанопоры: увеличение временного разрешения до предела диффузии. Датчики СКУД 2 , 1184–1190 (2017).
КАС
пабмед
Статья
Google Scholar
Hoenger, A., Pagès, J.M., Fourel, D. & Engel, A. Ориентация порина OmpF во внешней мембране Escherichia coli. Дж. Мол. биол. 233 , 400–413 (1993).
КАС
пабмед
Статья
Google Scholar
Данелон, К., Брандо, Т. и Винтерхальтер, М. Исследование ориентации реконструированных мальтопориновых каналов на уровне отдельных белков. J. Biol. хим. 278 , 35542–35551 (2003 г.).
КАС
пабмед
Статья
Google Scholar
Кульман Л., Винтерхальтер М. и Безруков С.М. Транспорт мальтодекстринов через мальтопорин: одноканальное исследование. Биофиз. J. 82 , 803–812 (2002).
Биофиз. J. 82 , 803–812 (2002).
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Баджадж, Х. и др. . Бактериальные порины внешней мембраны как электростатические наноситы: изучение правил транспорта малых полярных молекул. ACS Nano 11 , 5465–5473 (2017).
КАС
пабмед
Статья
Google Scholar
Красильников О.В., Мерзляк П.Г., Юлдашева Л.Н., Капистрано М.Ф. Белковая электрострикция: возможность упругой деформации α-гемолизинового канала приложенным полем. евро. Биофиз. J. 34 , 997–1006 (2005).
КАС
пабмед
Статья
Google Scholar
Аксиментьев А. и Шультен К. Визуализация α-гемолизина с помощью молекулярной динамики: ионная проводимость, осмотическая проницаемость и карта электростатического потенциала. Биофиз. J. 88 , 3745–3761 (2005).
Биофиз. J. 88 , 3745–3761 (2005).
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Кутцнер, К., Грубмюллер, Х., Де Гроот, Б.Л. и Захария, У. Вычислительная электрофизиология: молекулярная динамика проникновения и селективности ионных каналов в атомистических деталях. Биофиз. J. 101 , 809–817 (2011).
ОБЪЯВЛЕНИЕ
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Kutzner, C. и др. . Понимание функции ионных каналов с помощью компьютерного электрофизиологического моделирования. Биохим. Биофиз. Акта — биомембрана. 1858 , 1741–1752 (2016).
КАС
Статья
Google Scholar
Акоста-Гутьеррес, С., Скорчапино, М. А., Бодренко, И. и Чеккарелли, М. Фильтрация с помощью электрического поля: случай E. coli Porins. J. Phys. хим. лат. 6 , 1807–1812 (2015).
coli Porins. J. Phys. хим. лат. 6 , 1807–1812 (2015).
КАС
пабмед
Статья
Google Scholar
Gu, L.-Q., Cheley, S. & Bayley, H. Электроосмотическое усиление связывания нейтральной молекулы с трансмембранной порой. Проц. Натл. акад. науч. США 100 , 15498–15503 (2003 г.).
ОБЪЯВЛЕНИЕ
КАС
пабмед
Статья
Google Scholar
Ангелова М.И., Димитров Д.С. Электроформирование липосом. Фарадей Обсудить. хим. Soc 81 , 303–311 (1986).
КАС
Статья
Google Scholar
Хотц, Т. и др. . Идеализация записей ионного канала с помощью многоразрешающего фильтра сегментации скачков. IEEE Trans. Нанобиология 12 , 376–386 (2013).
ПабМед
Статья
Google Scholar

Джонс, Г., Уиллетт, П., Глен, Р. К., Лич, А. Р. и Тейлор, Р. Разработка и проверка генетического алгоритма для гибкой стыковки. Дж. Мол. биол. 267 , 727–748 (1997).
КАС
пабмед
Статья
Google Scholar
Морли, С. Д. и Афшар, М. Проверка эмпирической функции оценки РНК-лиганда для быстрой гибкой стыковки с использованием RiboDock. Дж. Вычисл. Помощь. Мол. Дес. 18 , 189–208 (2004).
ОБЪЯВЛЕНИЕ
КАС
пабмед
Статья
Google Scholar
Баррил Х., Хаббард Р. Э.и Морли, С. Д. Виртуальный скрининг при открытии лекарств на основе структуры. Mini Rev. Med. хим. 4 , 779–791 (2004).
КАС
пабмед
Google Scholar
Mulay, S.R. и др. . ProtSqueeze: Простой и эффективный автоматизированный инструмент для моделирования мембранных белков. J. Chem. Инф. Модель. 47 , 1986–1994 (2016).
J. Chem. Инф. Модель. 47 , 1986–1994 (2016).
Google Scholar
Wolf, M.G., Hoefling, M., Aponte-SantamaríA, C., Grubmüller, H. & Groenhof, G. G-membed: Эффективная вставка мембранного белка в уравновешенный липидный бислой с минимальным возмущением. Дж. Вычисл. хим. 31 , 2169–2174 (2010).
КАС
пабмед
Статья
Google Scholar
Линдорф-Ларсен, К. и др. . Улучшенные торсионные потенциалы боковой цепи для силового поля белка Amber ff99SB. Структура белков. Функц. Биоинформа. 78 , 1950–1958 (2010).
КАС
Google Scholar
Cordomí, A., Caltabiano, G. & Pardo, L. Моделирование мембранных белков с использованием силового поля AMBER и параметров липидов Бергера. J. Chem. Теория вычисл. 8 , 948–958 (2012).
ПабМед
Статья
Google Scholar

Финстра, К. А., Хесс, Б. и Берендсен, Х. Дж. К. Повышение эффективности крупномасштабного моделирования молекулярной динамики систем, богатых водородом. Дж. Вычисл. хим. 20 , 786–798 (1999).
КАС
Статья
Google Scholar
Lindahl, E. & Edholm, O. Мезоскопические неровности и флуктуации толщины липидных бислоев на основе моделирования молекулярной динамики. Биофиз. J. 79 , 426–433 (2000).
ОБЪЯВЛЕНИЕ
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Mark, P. & Nilsson, L. Структура и динамика моделей воды TIP3P, SPC и SPC/E при 298 K. J. Phys. хим. А 105 , 9954–9960 (2001).
КАС
Статья
Google Scholar
Joung, I. S. & Cheatham, T. E. Определение параметров щелочных и галогенидных одновалентных ионов для использования в явно сольватированных биомолекулярных моделированиях. J. Phys. хим. B 112 , 9020–9041 (2008 г.).
J. Phys. хим. B 112 , 9020–9041 (2008 г.).
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Ван, Дж., Вольф, Р. М., Колдуэлл, Дж. В., Коллман, П. А. и Кейс, Д. А. Разработка и испытание силового поля общего янтаря. Дж. Вычисл. хим. 25 , 1157–1174 (2004).
КАС
пабмед
Статья
Google Scholar
Wang, J., Cieplak, P. & Kollman, P. A. Насколько хорошо модель ограниченного электростатического потенциала (RESP) работает при расчете конформационной энергии органических и биологических молекул? Дж. Вычисл. хим. 21 , 1049–1074 (2000).
КАС
Статья
Google Scholar
Кейс и др. . ЯНТАРНЫЙ 12 . (Калифорнийский университет, Сан-Франциско, 2012 г.).
Авраам, М. Дж. и др. . Gromacs: Высокопроизводительное молекулярное моделирование с помощью многоуровневого параллелизма от ноутбуков до суперкомпьютеров. SoftwareX 1–2 , 19–25 (2015).
. Gromacs: Высокопроизводительное молекулярное моделирование с помощью многоуровневого параллелизма от ноутбуков до суперкомпьютеров. SoftwareX 1–2 , 19–25 (2015).
ОБЪЯВЛЕНИЕ
Статья
Google Scholar
Бусси, Г., Донадио, Д. и Парринелло, М. Каноническая выборка посредством масштабирования скорости. J. Chem. Физ . 126 (2007).
Берендсен, Х.Дж. К., Постма, Дж. П. М., ван Гюнстерен, В. Ф., ДиНола, А. и Хаак, Дж. Р. Молекулярная динамика с подключением к внешней ванне. J. Chem. физ. 81 , 3684–3690 (1984).
ОБЪЯВЛЕНИЕ
КАС
Статья
Google Scholar
Дарден Т., Йорк Д. и Педерсен Л. Сетка частиц Эвальд: метод N log(N) для сумм Эвальда в больших системах. J. Chem. физ. 98 , 10089 (1993).
ОБЪЯВЛЕНИЕ
КАС
Статья
Google Scholar

Хесс, Б., Беккер, Х., Берендсен, Х. Дж. К. и Фраайе, Дж. Г. Э. М. LINCS: Решатель линейных ограничений для молекулярного моделирования. Дж. Вычисл. хим. 18 , 1463–1472 (1997).
КАС
Статья
Google Scholar
Miyamoto, S. & Kollman, P. A. SETTLE: аналитическая версия алгоритма SHAKE and RATTLE для моделей жесткой воды. Дж. Вычисл. хим. 13 , 952–962 (1992).
КАС
Статья
Google Scholar
Мишо-Агравал, Н., Деннинг, Э. Дж., Вульф, Т. Б. и Бекштейн, О. MDAnalysis: набор инструментов для анализа моделирования молекулярной динамики. Дж. Вычисл. хим. 32 , 2319–2327 (2011).
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
McGibbon, R. T. и др. .MDTraj: современная открытая библиотека для анализа траекторий молекулярной динамики. Биофиз. J. 109 , 1528–1532 (2015).
Биофиз. J. 109 , 1528–1532 (2015).
ОБЪЯВЛЕНИЕ
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
Страница не найдена — ScienceDirect
Пандемия COVID-19 и глобальные изменения окружающей среды: новые потребности в исследованиях
Environment International, том 146, январь 2021 г., 106272.
Роберт Баруки, Манолис Кожевинас, […] Паоло Винеис
Исследования по количественной оценке риска изменения климата в городских масштабах: обзор недавнего прогресса и перспективы будущего направления
Обзоры возобновляемых и устойчивых источников энергии,
Том 135, январь 2021 г., 110415
Бин Йе, Цзинцзин Цзян, Цзюньго Лю, И Чжэн, Нань Чжоу
Воздействие изменения климата на экосистемы водно-болотных угодий: критический обзор экспериментальных водно-болотных угодий
Журнал экологического менеджмента,
Том 286, 15 мая 2021 г. , 112160
, 112160
Шокуфе Салими, Сухад А.А.А.Н. Альмуктар, Миклас Шольц
Обзор воздействия изменения климата на общество в Китае
Достижения в области исследований изменения климата,
Том 12, Выпуск 2, апрель 2021 г., страницы 210-223
Юн-Цзянь Дин, Чен-Ю Ли, […] Зенг-Ру Ван
Восприятие общественностью изменения климата и готовности к стихийным бедствиям: данные из Филиппин
2020
Винченцо Боллеттино, Тилли Алкайна-Стивенса, Манаси Шарма, Филип Дай, Фуонг Фама, Патрик Винк
Воздействие бытовой техники на окружающую среду в Европе и сценарии его снижения
Журнал чистого производства,
Том 267, 10 сентября 2020 г., 121952
Роланд Хишир, Франческа Реале, Валентина Кастеллани, Серенелла Сала
Влияние глобального потепления на смертность апрель 2021 г.
Раннее развитие человека,
Том 155, апрель 2021 г., 105222
Джин Кальеха-Агиус, Кэтлин Инглэнд, Невилл Кальеха
Понимание и противодействие мотивированным корням отрицания изменения климата
Текущее мнение об экологической устойчивости,
Том 42, февраль 2020 г. , страницы 60-64
, страницы 60-64
Габриэль Вонг-Пароди, Ирина Фейгина
Это начинается дома? Климатическая политика, направленная на потребление домохозяйствами и поведенческие решения, является ключом к низкоуглеродному будущему
Энергетические исследования и социальные науки
Том 52, июнь 2019 г., страницы 144–158.
Гилен Дюбуа, Бенджамин Совакул, […] Райнер Зауэрборн
Трансформация изменения климата: определение и типология для принятия решений в городской среде
Устойчивые города и общество,
Том 70, июль 2021 г., 102890
Анна С. Хурлиманн, Саре Мусави, Джеффри Р. Браун
«Глобальное потепление» против «изменения климата»: воспроизведение связи между политической самоидентификацией, формулировкой вопроса и экологическими убеждениями.
Журнал экологической психологии,
Том 69, июнь 2020 г., 101413
Алистер Рэймонд Брайс Суттер, Рене Мыттус
.
