▶▷▶ гдз история 6 класс рабочая тетрадь крючкова 2016 ответы
▶▷▶ гдз история 6 класс рабочая тетрадь крючкова 2016 ответы
| Интерфейс | Русский/Английский |
| Тип лицензия | Free |
| Кол-во просмотров | 257 |
| Кол-во загрузок | 132 раз |
| Обновление: | 10-11-2018 |
гдз история 6 класс рабочая тетрадь крючкова 2016 ответы — Yahoo Search Results Yahoo Web Search Sign in Mail Go to Mail» data-nosubject=»[No Subject]» data-timestamp=’short’ Help Account Info Yahoo Home Settings Home News Mail Finance Tumblr Weather Sports Messenger Settings Yahoo Search query Web Images Video News Local Answers Shopping Recipes Sports Finance Dictionary More Anytime Past day Past week Past month Anytime Get beautiful photos on every new browser window Download ГДЗ по истории 6 класс рабочая тетрадь Крючкова gdz-putinainfo 6 класс История ГДЗ » 6 класс » История ГДЗ готовые домашние задания к рабочей тетради Крючкова 6 класс по истории средних веков 2016 1 и 2 часть ФГОС от Путина ГДЗ рабочая тетрадь по истории 6 класс Крючкова botanamnet … 6 класс История Подробный решебник ГДЗ к рабочей тетради по истории 6 класс Крючкова ЕА 2013, онлайн ответы на домашнюю работу Решебник (ГДЗ) к рабочей тетради ЕА Крючкова, Всеобщая 30schoolru/ 6 -klass/istoriya/reshebnik-gdz-k-rabochej Cached История средних веков, 6 класс 17-ое издание, 2014 год Готовые ответы к рабочей тетради по Истории средних веков , автор ЕА Крючкова (2014 год 17е издание) Гдз История 6 Класс Рабочая Тетрадь Крючкова 2016 Ответы — Image Results More Гдз История 6 Класс Рабочая Тетрадь Крючкова 2016 Ответы images ГДЗ по Истории за 6 класс рабочая тетрадь ЕА Крючкова megareshebaru/gdz/istorija/ 6 -klass/rabochaya Cached ГДЗ по истории 6 класс рабочая тетрадь ЕА Крючкова автор: ЕА Крючкова Подробный решебник ( ГДЗ ) по Истории для 6 класса рабочая тетрадь , ГДЗ от Путина 6 класс История gdz-putinainfo/ 6 -klass/istoriya- 6 Cached ГДЗ от Путина 6 класс История решебники учебников и рабочих тетрадей по истории за 6 класс онлайн Данные гдз книг и тетрадей помогут вам проверить выполненное домашние задание ГДЗ 6 класс История ЯГДЗ — yagdzcom yagdzcom/ 6 -klass/istoriya- 6 Cached ЯГДЗ 6 класс История готовые домашние задания Ответы на задания и вопросы из учебников и рабочих тетрадей по истории за 6 класс (Агибалова, Донской, Ведюшкин, Данилов, Косулина, Пчелов, Пономарев) ГДЗ по истории 6 класс рабочая тетрадь Крючкова История yagdzcom 6 класс История ГДЗ по истории 6 класс рабочая тетрадь Крючкова ГДЗ решебник к рабочей тетради по истории 6 класс Крючкова История средних веков ФГОС 18 издание ГДЗ по истории 6 класс рабочая тетрадь Крючкова otlgdzcom 6 КЛАСС ГДЗ на Отлично » 6 КЛАСС » ГДЗ по истории для 6 класса ГДЗ по истории 6 класс рабочая тетрадь Крючкова История России, изучаемая в 6 классе это часть всемирной истории человечества Рабочая тетрадь по истории России 6 класс Артасов, Данилов 2016 newgdzcom/istoriya-5-11-klass-uchebniki-onlajn/istoriya Артемов, Соколова тетрадь для проектов и творческих работ 6 класс всеобщая история средних веков 2017 Рабочая тетрадь по всеобщей истории средних веков 6 класс Ведюшкин, Крючкова 2014 ГДЗ по истории за 6 класс решебники и рабочие тетради botanamnet Решебники 6 класс ГДЗ рабочая тетрадь по истории 6 класс Данилов Косулина ГДЗ рабочая тетрадь по истории 6 класс Крючкова 2015- 2016 Promotional Results For You Free Download | Mozilla Firefox ® Web Browser wwwmozillaorg Download Firefox — the faster, smarter, easier way to browse the web and all of Yahoo 1 2 3 4 5 Next 25,900 results Settings Help Suggestions Privacy (Updated) Terms (Updated) Advertise About ads About this page Powered by Bing™
- составленный как сборник ГДЗ к рабочей тетради педагога Крючковой Елены Александровны
- существенно облегчает участь шестиклассников Пособие составлено учителями
- включая контурные карты
«Города
«Люди и события Средневековья»
- Пчелов
- Данилов 2016 newgdzcom/istoriya-5-11-klass-uchebniki-onlajn/istoriya Артемов
- Данилов
Яндекс Яндекс Найти Поиск Поиск Картинки Видео Карты Маркет Новости ТВ онлайн Музыка Переводчик Диск Почта Коллекции Все Ещё Дополнительная информация о запросе Показаны результаты для Нижнего Новгорода Москва 1 ГДЗ по истории 6 класс рабочая тетрадь Крючкова GDZ-Putinainfo › 6 класс › История › …-tetrad-kryuchkova Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте ГДЗ готовые домашние задания к рабочей тетради Крючкова 6 класс по истории средних веков 2016 1 и 2 часть ФГОС от Путина Решебник ( ответы на вопросы и задания) 2 ГДЗ по истории 6 класс рабочая тетрадь Крючкова eurokiorg › gdz…istoriya/6_klass…tetrad…kruchkova… Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте ГДЗ рабочая тетрадь по истории средних веков 6 класс Крючкова Просвещение История средних веков, изучаемая в 6 Решебник, составленный как сборник ГДЗ к рабочей тетради педагога Крючковой Елены Александровны, существенно Читать ещё ГДЗ рабочая тетрадь по истории средних веков 6 класс Крючкова Просвещение История средних веков, изучаемая в 6 классе — интереснейшая наука, но осмысление давно минувших событий порою непонятно нынешним школьникам, заполнение контурных карт представляется им тяжким трудом Учебник ФГОС скрупулезно штудировать способны немногие ученики Решебник, составленный как сборник ГДЗ к рабочей тетради педагога Крючковой Елены Александровны, существенно облегчает участь шестиклассников Пособие составлено учителями, высокая точность ответов гарантирована Скрыть 3 ГДЗ по истории 6 класс ( рабочая тетрадь ) Крючкова gdzmaniacom › gdz…istoriya-6…kryuchkova…tetradhtml Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Автор: ЕА Крючкова Мы добавили качественный решебник к рабочей тетради автора Крючкова по предмету » История Средних веков» за 6 класс Данный сборник содержит готовые онлайн ответы на все вопросы и задания тетради , который поможет шестик Читать ещё Автор: ЕА Крючкова Мы добавили качественный решебник к рабочей тетради автора Крючкова по предмету » История Средних веков» за 6 класс Данный сборник содержит готовые онлайн ответы на все вопросы и задания тетради , который поможет шестиклассникам быстро и уверенно справиться с домашней и школьной работой Стоит отметить, что эти ответы в первую очередь созданы для родителей школьников, чтобы они могли без проблем проконтролировать уроки своего ученика по истории в шестом классе Cмотреть тут: Агибалова Донской (учебник) Скрыть 4 ГДЗ по истории 6 класс Крючкова ( рабочая тетрадь ) gdzroomorg › istoriya…klass-kryuchkova…tetradhtml Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Онлайн ГДЗ к рабочей тетради Крючковой по истории 6 класса Ниже вы найдете ответы на все задания тетради , включая контурные карты, кроссворды, разделы «Теперь вы это знаете», «Люди и события Средневековья», «Эти слова пришли к нам из Средневековья», «Города, страны, континенты» Читать ещё Онлайн ГДЗ к рабочей тетради Крючковой по истории 6 класса Ниже вы найдете ответы на все задания тетради , включая контурные карты, кроссворды, разделы «Теперь вы это знаете», «Люди и события Средневековья», «Эти слова пришли к нам из Средневековья», «Города, страны, континенты», «Итоговые задания » Приятного пользования вам желает портал gdzroomorg Скрыть 5 ГДЗ по истории 6 класс Крючкова рабочая тетрадь GDZme › 6 класс › История › Крючкова рабочая тетрадь Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте ГДЗ по истории 6 класс к рабочей тетради Крючкова , онлайн ответы из решебника Рабочая тетрадь по истории за 6 класс автора Крючковой ЕА 2013 года издания Сборник состоит из готовых решений на многочисленные упражнения разного вида и уровня сложности Выполненные задания Читать ещё ГДЗ по истории 6 класс к рабочей тетради Крючкова , онлайн ответы из решебника Рабочая тетрадь по истории за 6 класс автора Крючковой ЕА 2013 года издания Сборник состоит из готовых решений на многочисленные упражнения разного вида и уровня сложности Выполненные задания предусмотрены для учащихся с разным показателем знаний, включая и самый низкий Скрыть 6 Ответы ГДЗ рабочая тетрадь по истории Крючкова dourokovru › 6-klass/istoriya…tetrad…kryuchkova-6… Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Главная › 6 класс › История Рабочая тетрадь по истории Крючкова 6 класс Содержание рабочей тетради §1 Образование варварских Читать ещё Главная › 6 класс › История Рабочая тетрадь по истории Крючкова 6 класс Содержание рабочей тетради §1 Образование варварских королевств Государство франков в VI – VIII стр 3-7 §2 Христианская церковь в раннее Средневековье стр 8-10 §3 Возникновение и распад империи Карла Великого стр 10-12 Скрыть 7 Рабочая тетрадь по истории 6 класс Крючкова tetrabru › 6-klass/istoriya…tetrad…kryuchkovahtml Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Главная › 6 класс › История › Рабочая тетрадь по истории 6 класс Крючкова Государство франков в VI – VIII §2 «Христианская церковь в раннее Средневековье» Читать ещё Главная › 6 класс › История › Рабочая тетрадь по истории 6 класс Крючкова Рабочая тетрадь по истории 6 класс Крючкова Авторы: Крючкова Е А Государство франков в VI – VIII §2 «Христианская церковь в раннее Средневековье» §3 «Возникновение и распад империи Карла Великого» Скрыть 8 ГДЗ решебник по истории 6 класс рабочая тетрадь gdzpopme › 6-klass/istoriya…tetrad-kryuchkovahtml Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Мы предлагаем Вам ответы к рабочей тетради Крючкова 6 класс по истории 2013-2015 1 и 2 часть Вы можете смотреть и читать 9 ГДЗ история 6 класс рабочая тетрадь Крючкова 2016 ответы — смотрите картинки ЯндексКартинки › гдз история 6 класс рабочая тетрадь крючкова 2016 Пожаловаться Информация о сайте Ещё картинки 10 ГДЗ по истории 6 класс рабочая тетрадь Крючкова otlGDZonline › 6…istoriya-6…tetrad-kryuchkovahtml Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте История России, изучаемая в 6 классе это часть всемирной истории человечества Учащимся предлагают спланировать пути достижения учебных задач, самостоятельно определить и скорректировать их ГДЗ рабочая тетрадь по истории 6 класс Крючкова torgunet › gdz/6-klass/istoriya/kryuchkova-122/ Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Публикуем вам ГДЗ к рабочей тетради по истории за шестой класс авторов Крючкова ЕА 2014 года издания Вы можете читать и смотреть готовые домашние задания по истории на смартфоне, компьютере или телефоне Читать ещё Публикуем вам ГДЗ к рабочей тетради по истории за шестой класс авторов Крючкова ЕА 2014 года издания Вы можете читать и смотреть готовые домашние задания по истории на смартфоне, компьютере или телефоне Ответы Ответы на задания по истории за шестой класс к рабочей тетради Крючкова ЕА Скрыть Вместе с « гдз история 6 класс рабочая тетрадь крючкова 2016 ответы » ищут: гдз история 6 класс рабочая тетрадь чернова гдз по истории 6 класс рабочая тетрадь чернова история средних веков гдз история 6 класс контурные карты средние века гдз история 6 класс учебник гдз по математике 6 класс виленкин гдз по русскому языку 6 класс ладыженская как византийские историки объясняли ход истории и причины событий 1 2 3 4 5 дальше Браузер Все новые вкладки с анимированным фоном 0+ Установить
ГДЗ по Истории 6 класс Проверочные и контрольные работы Крючкова
С «ГДЗ по истории Средних веков за 6 класс к проверочным и контрольным работам Крючковой» нет ничего невозможного. И это не просто слова. К примеру, раньше ученик часто пропускал школу по неуважительным причинам, не серьезно относился к данной дисциплине. В результате этого, у него накопилось очень много пробелов и не обошлось без отрицательных оценок. Чтобы все исправить, нужно вооружиться надежным онлайн-пособием, коим является этот замечательный сборник правильных ответов и решенных номеров. Этот предмет хоть и интересный, но он не такой легкий, как многим может показаться на первый взгляд. На каждом уроке ребятам приходится усваивать очень много новой информации. К примеру, в этом году они будут проходить следующие темы:
И это не просто слова. К примеру, раньше ученик часто пропускал школу по неуважительным причинам, не серьезно относился к данной дисциплине. В результате этого, у него накопилось очень много пробелов и не обошлось без отрицательных оценок. Чтобы все исправить, нужно вооружиться надежным онлайн-пособием, коим является этот замечательный сборник правильных ответов и решенных номеров. Этот предмет хоть и интересный, но он не такой легкий, как многим может показаться на первый взгляд. На каждом уроке ребятам приходится усваивать очень много новой информации. К примеру, в этом году они будут проходить следующие темы:
- Восточные славяне в VII-IX вв.
- Большая феодальная война.
- Правление Ивана III. Внешняя политика.
- Приемники Золотой Орды.
- Московская Русь.
Для чего нужен решебник по истории Средних веков за 6 класс к проверочным и контрольным работам Крючковой
Разделы очень объемные. За один или два урока учитель просто физически не успевает ни объяснить параграфы, ни поработать с каждым индивидуально. Ученикам приходится самим выкручиваться из сложившейся ситуации. Чтобы ребята не прибегали к помощи сомнительных источников информации, им следует сразу же вооружиться справочником с подробным описанием решений всех заданий. Многие дети часто заглядывают в эту книгу, чтобы:
За один или два урока учитель просто физически не успевает ни объяснить параграфы, ни поработать с каждым индивидуально. Ученикам приходится самим выкручиваться из сложившейся ситуации. Чтобы ребята не прибегали к помощи сомнительных источников информации, им следует сразу же вооружиться справочником с подробным описанием решений всех заданий. Многие дети часто заглядывают в эту книгу, чтобы:
- проверить, верно ли они ответили на тот или иной вопрос;
- расшифровать условие упражнения;
- понять принцип его решения.
«ГДЗ по истории Средних веков за 6 класс к проверочным и контрольным работам Крючковой Е. А. (Просвещение)» прекрасно справится с ролью персонального консультанта. С ним можно пройти через все испытания. Некоторые школьники успели убедиться в эффективности данного справочника. А другим только предстоит с ним познакомиться. Сборник верных ответов является своего рода учебно-вспомогательной литературой.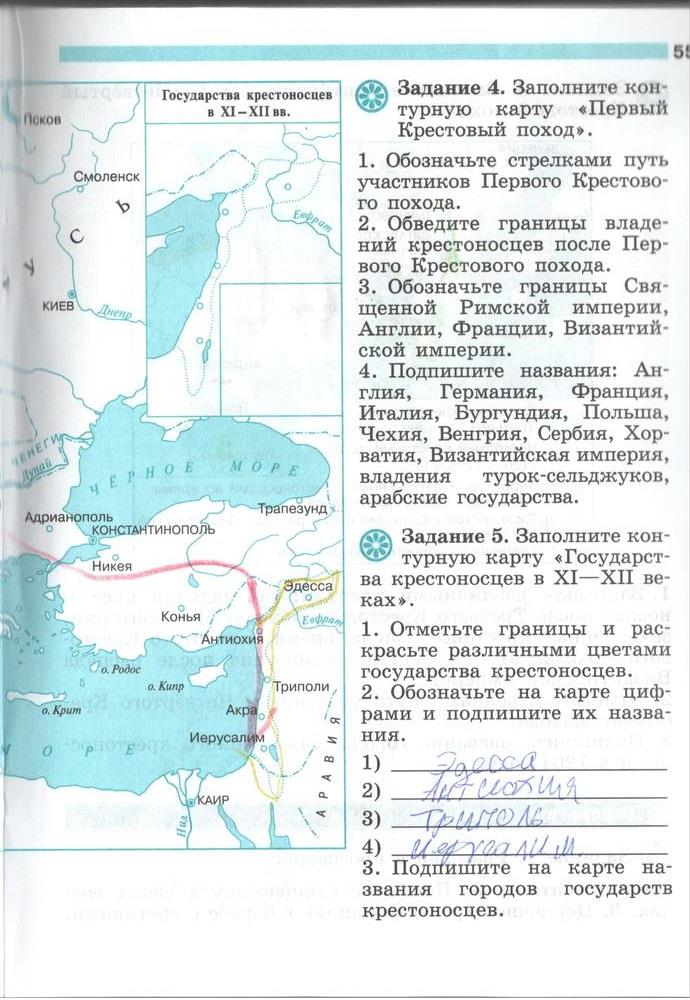 Он был разработан специалистом высокой квалификации специально для того, чтобы с его помощью шестиклассники смогли облегчить процесс обучения в школе и не потерять интерес к данной дисциплине. Плюс ко всему, это пособие избавит ребят от необходимости часами просиживать за учебниками и дополнительными справочниками. Им удастся за считанные минуты выполнить домашнее задание, а еще более тщательно подготовиться к обычной самостоятельной, опросу в классе или же важному итоговому тесту.
Он был разработан специалистом высокой квалификации специально для того, чтобы с его помощью шестиклассники смогли облегчить процесс обучения в школе и не потерять интерес к данной дисциплине. Плюс ко всему, это пособие избавит ребят от необходимости часами просиживать за учебниками и дополнительными справочниками. Им удастся за считанные минуты выполнить домашнее задание, а еще более тщательно подготовиться к обычной самостоятельной, опросу в классе или же важному итоговому тесту.
ГДЗ к учебнику История Средних веков 6 класс Агибалов можно посмотреть
здесь.
ГДЗ История Средних веков 6 класс рабочая тетрадь Крючкова можно посмотреть
здесь.
ГДЗ к контурным картам История Средних веков за 6 класс Ведюшкин можно посмотреть
здесь.
ГДЗ к контрольно-измерительным материалам История Средних веков за 6 класс Волкова можно посмотреть
здесь.
ГДЗ к рабочей тетради по истории Средних веков за 6 класс Чернова можно посмотреть
здесь.
ГДЗ к рабочей тетради УУД по истории Средних веков за 6 класс Чернова можно посмотреть
здесь.
ГДЗ по Истории за 6 класс Рабочая тетрадь Е.А. Крючкова
История 6 класс
Е.А. Крючкова
рабочая тетрадь
Авторы: Е.
«Решебник по Истории 6 класс Рабочая тетрадь Крючкова (Просвещение)» станет надежным союзником ученику средней школы в изучении одной из сложнейших наук школьной программы.
ГДЗ призван помочь преодолеть все возникающие проблемы у школьников, а также позволит им максимально быстро и качественно разобраться в трудной теме. Основные плюсы ГДЗ:
- онлайн-размещение на сайте и круглосуточный доступ;
- поможет существенно сократить среднее время выполнения домашнего задания;
- позволит ученикам подтянуть оценки и приумножить имеющиеся знания;
- даст возможность школьникам проявить себя с лучшей стороны при ответе на уроке, а учитель обязательно отметит положительной оценкой такую тягу к учебе;
- несет в себе верные ответы на все упражнения.
Немаловажно и то, что учебник разработан в строгом соответствии со всеми требованиями федерального государственного образовательного стандарта, он рекомендован к изучению в школах министерством образования. Следовательно, номера в решебнике, которые полностью соответствуют основному изданию, отвечают всем требованиям учебного стандарта.
Следовательно, номера в решебнике, которые полностью соответствуют основному изданию, отвечают всем требованиям учебного стандарта.
Процесс изучения истории в средней школе
В этом году ребятам предстоит изучить следующие темы:
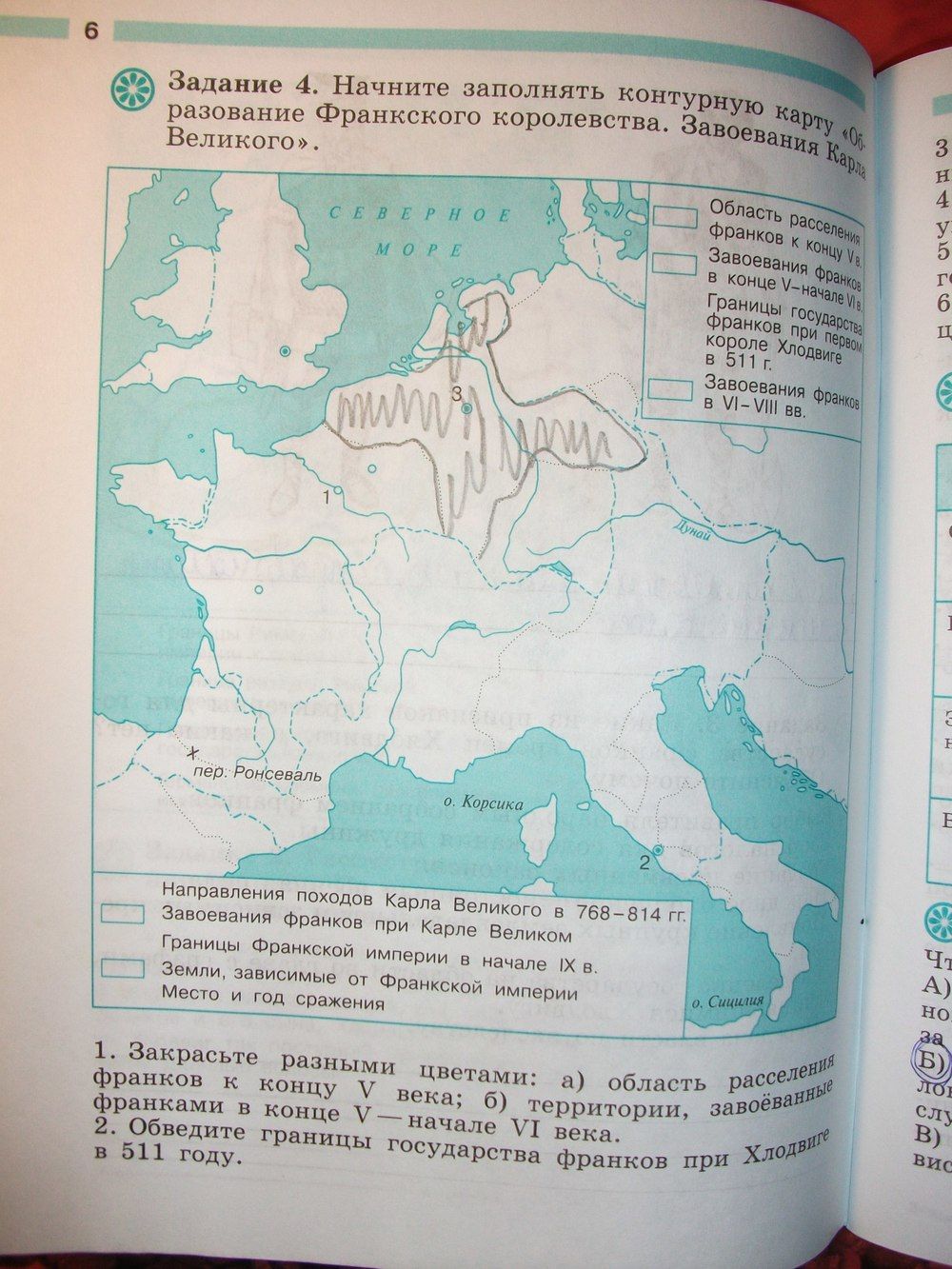
- великое переселение народов и знаменитые варварские королевства Западной Европы;
- закат Византийской империи, причины;
- завоевания норманнов и быт Англии в раннем Средневековье;
- предпосылки и развитие феодального общества в Западной Европе;
- папское могущество в священной Римской империи.
Чтобы на хорошем уровне овладеть материалом перечисленных разделов учебника, рекомендуется воспользоваться вспомогательным ресурсом – «ГДЗ по Истории 6 класс Рабочая тетрадь Крючкова Е.А. (Просвещение)». Он станет сертифицированным и грамотно составленным карманным помощником для школьника в учебе.
Рекомендации по работе с ГДЗ по истории за 6 класс от Крючковой
Для того, чтобы использование решебника принесло желаемый результат ученику шестого класса, стоит научиться с ним правильно работать, и полностью исключить бездумное списывание.
Мы выделили рекомендуемый порядок действий:
- прочитать теоретический материал по изучаемой теме;
- ответить на вопросы после текста и закрепить прочитанное;
- самостоятельно выполнить домашние задания;
- сделать самопроверку по верным ответам из решебника;
- произвести работу над ошибками.
Используя вышеуказанный комплекс по взаимодействию с пособием ГДЗ, школьник обязательно достигнет высоких результатов и получит массу положительных оценок от учителя.
ГДЗ ответы — учебники, рабочие тетради по Истории за 6 класс
Кто бы что ни говорил, а изучение своей культуры и наследия – очень важный период в жизни каждого школьника. Это требует не только интереса, но ещё и различной дополнительной информации. А на её поиск всегда уходит время. Трудно представить человека, который не был знаком с нашим прошлым, хотя бы косвенно. Однако можно углубить свои знания! Учебно-методический комплекс, который всегда доступен на нашем уникальном сервисе, не только подтянет по школьной программе, но и покажет каким был мир на всех этапах формирования.
Почему стоит выбрать онлайн-решебник по истории за 6 класс
Наши онлайн-гдз были разработаны не только для того, чтобы из него школьники просто переписывали верные ответы в чистовик. Данный решебник преследует более важные цели. Во-первых, благодаря ему дети учатся применять полученные знания в деле. Во-вторых, к изучению этого, и остальных предметов они подходят со всей серьезностью и ответственностью. Рабочая программа для нового учебного года направлена на изучение следующих тем:
- Белорусские города в составе Смоленской и Черниговской земель.
- Балтские соседи Руси на северо-западе.
- Внешняя опасность в первой половине XIII в.
- Возникновение городов и княжеств в Понёманье.
- Появление крестоносцев в Восточной Прибалтике.
- Монголо-татарское нашествие.
После каждого раздела ученикам предстоит ответить на ряд контрольных вопросов. Это поможет педагогу понять кто и как усвоил материал, а с кем еще следует немного поработать.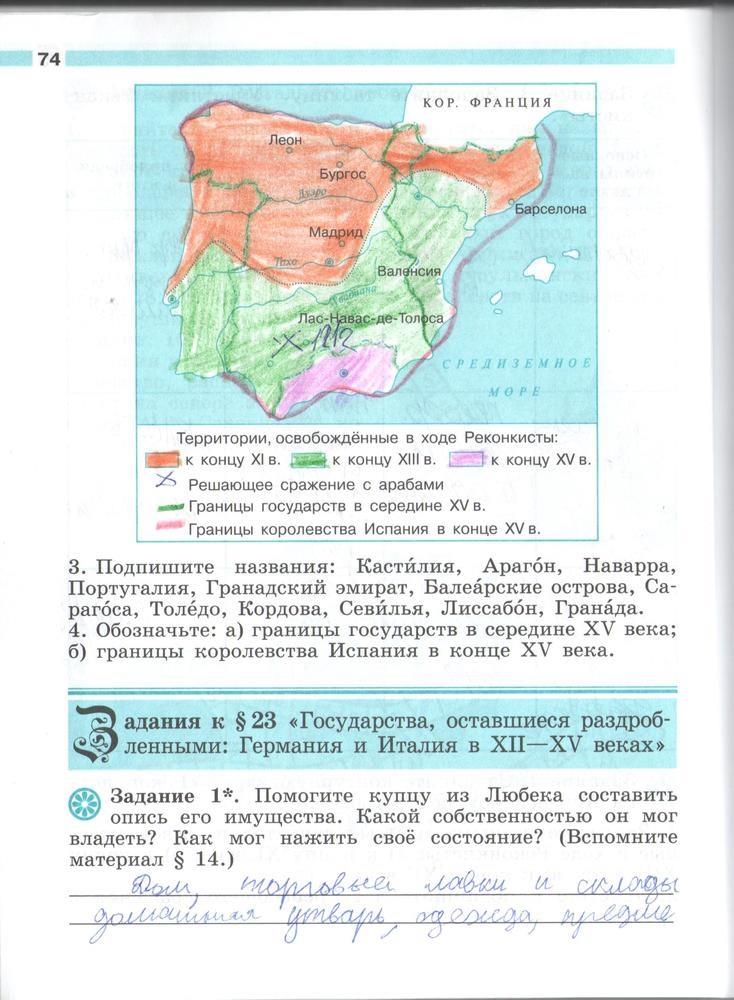 В любом случае оценки за подобные тесты очень важны. Чтобы получить желаемую отметку за работу, ребенок должен приходить на уроки хорошо подготовленным с выполненным домашним заданием. А если на его пути возникнут трудности, то поможет с ними справиться данный учебно-методический комплекс, разработанный высококвалифицированными специалистами.
В любом случае оценки за подобные тесты очень важны. Чтобы получить желаемую отметку за работу, ребенок должен приходить на уроки хорошо подготовленным с выполненным домашним заданием. А если на его пути возникнут трудности, то поможет с ними справиться данный учебно-методический комплекс, разработанный высококвалифицированными специалистами.
Практика с ГДЗ по истории для шестых классов
Работать с данным пособием ученики могут так, как им удобно. Но если ребенок серьезно отстал от программы, то ему следует какое-то время придерживаться определенного алгоритма. Для начала он должен сам выполнять заданные на дом упражнения. После этого он может сверять ответы, и проводить работу над ошибками. Эти два действия являются обязательными. Без них школьнику не удастся восполнить образовавшиеся пробелы в знаниях, и разобраться во всех тонкостях и нюансах новых тем. Каждое решенное задание сопровождается развернутыми авторскими комментариями. Это поможет шестикласснику понять, почему ответ получился именно таким, а не другим, и как авторы (А. А. Данилов, Л.Г. Косулина, Е. В. Агибалов, Г. М. Донской, Искровская Л.В., Федоров С.Е., Гурьянова Ю.В., Баранов П.А., Ермолаева Л.К., Лебедева И.М., Шейко Н.Г., Гурьянова Ю.В.) пришли к тем или иным выводам. Если вы тоже придерживаетесь взглядов, что решебник неэффективен для учебы в школе, значит, глубоко ошибаетесь. Да, дети привыкли пользоваться шпаргалками, не задумываясь о том, что такое списывание поможет лишь в сокращении времени, которое приходится тратить на выполнение домашней работы. Весь секрет в том, что нужно уходить от списывания и настроиться на серьёзное изучение материала. Небольшие рекомендации к использованию:
А. Данилов, Л.Г. Косулина, Е. В. Агибалов, Г. М. Донской, Искровская Л.В., Федоров С.Е., Гурьянова Ю.В., Баранов П.А., Ермолаева Л.К., Лебедева И.М., Шейко Н.Г., Гурьянова Ю.В.) пришли к тем или иным выводам. Если вы тоже придерживаетесь взглядов, что решебник неэффективен для учебы в школе, значит, глубоко ошибаетесь. Да, дети привыкли пользоваться шпаргалками, не задумываясь о том, что такое списывание поможет лишь в сокращении времени, которое приходится тратить на выполнение домашней работы. Весь секрет в том, что нужно уходить от списывания и настроиться на серьёзное изучение материала. Небольшие рекомендации к использованию:
- Перед ребенком незнакомое и тяжелое задание. Прочтите его внимательно, и постарайтесь решить самостоятельно.
- Если не выходит, обращайтесь к пособию, проанализируйте каждый шаг готового решения.
- В случае, если решить задачу получилось, необходимо сверить ответ, и исправить ошибки, которые возможно были допущены.
Если следовать инструкции, школьник не только освободит время для личного досуга, заработает хорошую оценку, но и существенно повысит уровень знаний.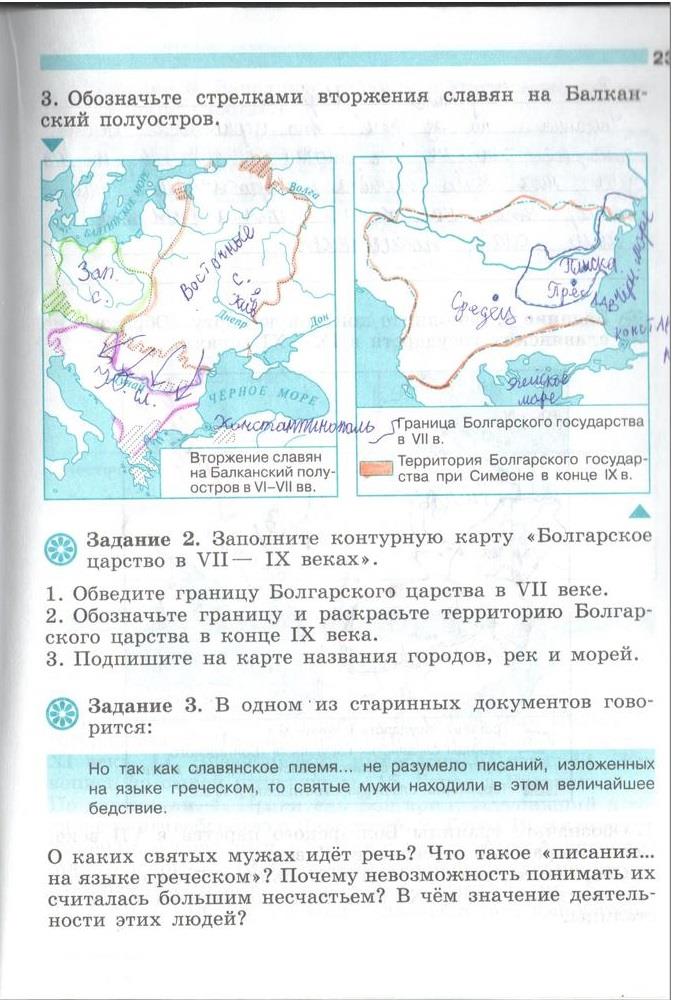 История откроется с новой неожиданной стороны. Делаем вывод, что представленный онлайн-справочник будет отличным помощником в учебе. При столкновении с похожим типом работ, не придётся тратить время на изучение вопроса, а можно сразу приступить к решению заданий.
История откроется с новой неожиданной стороны. Делаем вывод, что представленный онлайн-справочник будет отличным помощником в учебе. При столкновении с похожим типом работ, не придётся тратить время на изучение вопроса, а можно сразу приступить к решению заданий.
ГДЗ История 6 класс Крючкова
- История 6 класс
- Тип пособия: Рабочая тетрадь
- Авторы: Крючкова
- Издательство: «Просвещение»
Похожие ГДЗ История 6 класс
Задания: стр.3
Предыдущее
Следующее
Предыдущее
Следующее
История Средневековья – это реально интересный период, насыщенный увлекательными событиями, которые не всегда понятны и доступны для осознания многим учащимся шестых классов средней школы. Большинство ребят с огромным интересом смотрят исторические фильмы, некоторые зачитываются историческими романами, но изучать многочисленные даты и биографии великих личностей нравится далеко не всем. Отдельного внимания заслуживают картографические задания. Для качественного усвоения информации следует изучать разносторонние источники.
Для качественного усвоения информации следует изучать разносторонние источники.
Особенности использования решебника
Решебник к учебнику «История 6 класс Рабочая тетрадь Крючкова Просвещение» – верный помощник для подавляющего большинства учащихся шестых классов средней школы. Материал разработан педагогами, которые обладают большим опытом в данной сфере, по этой причине точность и правильность ответов гарантирована. Решебник пригодится, если нужно проверить готовое задание, разобраться в непростом упражнении или мало времени на выполнения домашнего задания.
В чем состоит преимущество решебника
Наличие такого пособия позволяет экономить силы и личное время родителей.
Многие родители в ГДЗ видят ряд положительных сторон:
- качественная, быстрая и точная проверка домашнего задания у ребенка;
- восстановление забытых знаний по предмету у детей;
- четкий контроль за школьником, как стимуляция учебного фактора.
Списывание исключается, так как с решебником необходимо правильно работать.
Как лучше использовать решебник
Взрослые имеют право спрятать в недоступное место пособие, однако грамотный ребенок извлечет исключительно пользу от готовых домашних заданий. Рекомендуется грамотно использовать решебник, а не просто механически списывать, однако, и такой разовый метод применения готовых домашних заданий принесет свои плоды – ребенок непроизвольно запомнит переписанные ответы. На ста двенадцати страницах решебника можно найти вопросы и ответы по всему курсу истории шестого класса:
- О рыцарях и короле Артуре.
- О культуре Византии.
- О столетней войне между Англией и Францией.
- О путешествиях Марко Поло.
- О достижениях Иоанна Гутенберга.
Нередко наличие Готовых Домашних Заданий к рабочей тетради по истории встречается и у педагогов.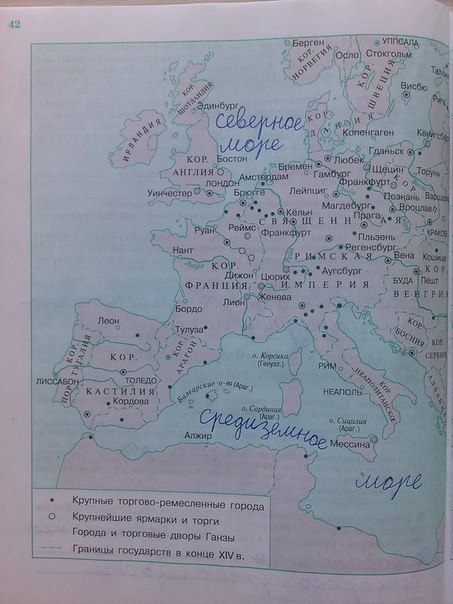 С помощью данного отличного пособия учителя могут быстро проверять работы учеников и качественно готовиться к урокам.
С помощью данного отличного пособия учителя могут быстро проверять работы учеников и качественно готовиться к урокам.
Последствия для комплексных исследований заболеваний и ассоциаций
Abstract
Накопление умеренно вредных миссенс-мутаций в индивидуальных геномах человека было предложено в качестве генетической основы для сложных заболеваний. Правдоподобие этой гипотезы зависит от количественных оценок распространенности умеренно вредных мутаций de novo и полиморфных вариантов у человека и от интенсивности селективного давления на них. Мы объединили анализ мутаций, вызывающих менделевские болезни человека, дивергенции между человеком и шимпанзе и систематических данных о генетической изменчивости человека и обнаружили, что ∼20% новых миссенс-мутаций у людей приводят к потере функции, тогда как ∼27% эффективно нейтральны. .Таким образом, оставшиеся 53% новых миссенс-мутаций имеют умеренно вредные последствия. Эти мутации приводят к появлению множества низкочастотных вредных аллельных вариантов в человеческой популяции, о чем свидетельствует новый набор данных о 37 генах, секвенированных в более чем 1500 отдельных хромосомах человека. Удивительно, но до 70% низкочастотных миссенс-аллелей умеренно вредны и связаны с гетерозиготной потерей приспособленности в диапазоне 0,001–0,003. Таким образом, низкая частота аллеля варианта аминокислоты сама по себе может служить предиктором его функциональной значимости.В нескольких недавних исследованиях сообщается о значительном избытке редких миссенс-вариантов в генах-кандидатах или путях у индивидуумов с экстремальными значениями количественных фенотипов. Эти исследования вряд ли дали бы результаты, если бы большинство редких вариантов были нейтральными или если бы редкие варианты не вносили значительный вклад в генетический компонент фенотипического наследования. Наши результаты обеспечивают обоснование этих типов исследований ассоциации генов-кандидатов (путей) и подразумевают, что баланс мутации-отбора может быть возможным эволюционным механизмом, лежащим в основе некоторых распространенных заболеваний.
Удивительно, но до 70% низкочастотных миссенс-аллелей умеренно вредны и связаны с гетерозиготной потерей приспособленности в диапазоне 0,001–0,003. Таким образом, низкая частота аллеля варианта аминокислоты сама по себе может служить предиктором его функциональной значимости.В нескольких недавних исследованиях сообщается о значительном избытке редких миссенс-вариантов в генах-кандидатах или путях у индивидуумов с экстремальными значениями количественных фенотипов. Эти исследования вряд ли дали бы результаты, если бы большинство редких вариантов были нейтральными или если бы редкие варианты не вносили значительный вклад в генетический компонент фенотипического наследования. Наши результаты обеспечивают обоснование этих типов исследований ассоциации генов-кандидатов (путей) и подразумевают, что баланс мутации-отбора может быть возможным эволюционным механизмом, лежащим в основе некоторых распространенных заболеваний.
Многие распространенные заболевания человека имеют сильный наследственный компонент. Хотя современная генетика человека добилась невероятного успеха в определении генетических причин редких менделевских заболеваний, комплексные заболевания оказались более сложной проблемой. 1 , 2 Генетическая изменчивость, влияющая на восприимчивость человека к большинству таких заболеваний, до сих пор в значительной степени неизвестна. Особый интерес представляют миссенс-мутации, поскольку считается, что многие из них обладают немаргинальными функциональными эффектами. 1 Влияние миссенс-мутации на молекулярную функцию, фенотип и приспособленность организма может быть чрезвычайно разнообразным. Миссенс-мутация может привести к летальному исходу или вызвать тяжелую менделевскую болезнь; в качестве альтернативы, он может быть слегка вредным, фактически нейтральным или полезным. Знание относительной частоты этих типов мутаций и их вклада в популяционную генетическую изменчивость важно для понимания эволюционного фона распространенных заболеваний и может помочь в планировании генетических исследований человека.
Хотя современная генетика человека добилась невероятного успеха в определении генетических причин редких менделевских заболеваний, комплексные заболевания оказались более сложной проблемой. 1 , 2 Генетическая изменчивость, влияющая на восприимчивость человека к большинству таких заболеваний, до сих пор в значительной степени неизвестна. Особый интерес представляют миссенс-мутации, поскольку считается, что многие из них обладают немаргинальными функциональными эффектами. 1 Влияние миссенс-мутации на молекулярную функцию, фенотип и приспособленность организма может быть чрезвычайно разнообразным. Миссенс-мутация может привести к летальному исходу или вызвать тяжелую менделевскую болезнь; в качестве альтернативы, он может быть слегка вредным, фактически нейтральным или полезным. Знание относительной частоты этих типов мутаций и их вклада в популяционную генетическую изменчивость важно для понимания эволюционного фона распространенных заболеваний и может помочь в планировании генетических исследований человека.
Воздействие миссенс-мутации на организм всегда многогранно и может рассматриваться с разных точек зрения — биохимической, медицинской и эволюционной. Взаимосвязь между эффектами замещения аминокислот на активность белка, здоровье человека и эволюционную приспособленность человека нетривиальна. Мутация, которая повреждает структуру белка, не обязательно ведет к обнаруживаемому фенотипу болезни человека, а мутация, которая предрасполагает человека к болезни, не обязательно вредна для эволюции.Фиксация полностью дезактивирующих мутаций (псевдогенезация), по-видимому, была обычным явлением в ходе недавней эволюции человека, 3 указывающая на то, что мутация, устраняющая активность белка, не обязательно подвергается очищающему отбору. Замещения, приводящие к аномальной функции гемоглобина, которые вызывают серповидно-клеточную анемию, очевидно, негативны как с биохимической, так и с медицинской точек зрения. Тем не менее их нельзя считать негативными с эволюционной точки зрения, потому что балансирующий отбор привел к их высокой частоте во многих частях мира в результате резистентности к малярии у гетерозигот. Чтобы четко различать различные аспекты негативных мутаций, мы используем термин «повреждающие» для обозначения мутации, снижающей активность белка, термин «вредные» для обозначения мутаций, предрасполагающих человека к заболеванию, и термин «вредные». для обозначения мутации, подвергшейся очищающему отбору.
Чтобы четко различать различные аспекты негативных мутаций, мы используем термин «повреждающие» для обозначения мутации, снижающей активность белка, термин «вредные» для обозначения мутаций, предрасполагающих человека к заболеванию, и термин «вредные». для обозначения мутации, подвергшейся очищающему отбору.
Высокий уровень заболеваемости многими сложными заболеваниями предполагает, что в человеческой популяции должна присутствовать удивительно высокая кумулятивная частота вредных для здоровья вариантов.Остается неясным, почему такие полиморфизмы могут сохраняться, не устраняясь очищающим отбором. В настоящее время существуют два основных направления рассуждений, объясняющих этот кажущийся парадокс. Первый рассматривает различные сложные эволюционные сценарии и рассматривает положительный или уравновешивающий отбор как основную силу, которая может привести к высокой частоте вредных для здоровья мутаций. Вторая линия рассуждений постулирует высокую частоту мутаций как основной фактор, определяющий кумулятивную частоту вредных полиморфизмов в популяции.
Первая гипотеза утверждает, что большинство полиморфизмов, предрасполагающих человека к сложному заболеванию, хотя и вредны с медицинской точки зрения в настоящее время, не были вредны для эволюции. Есть несколько возможных феноменов, которые могут помочь полиморфному варианту, придающему восприимчивость к фенотипу болезни, избежать очищающего отбора. Один из них — позднее начало заболевания, когда пагубные фенотипические последствия мутации наступают после репродуктивного возраста и, таким образом, не влияют на численность потомства особи.Несколько более ранних исследований, однако, показали, что это явление вряд ли будет обычным явлением. Эти исследования предполагают, что большинство мутаций, влияющих на фенотипы в пожилом возрасте, также имеют небольшой плейотропный эффект в начале жизни. 4 , 5 Изменение направления отбора — еще один механизм, который может объяснить, как вредные в настоящее время мутации могли избежать очищающего отбора. 6 Человеческий образ жизни, окружающая среда и питание резко изменились, и некоторые мутации, которые были нейтральными или даже полезными в обществах охотников-собирателей десятки тысяч лет назад, могут стать вредными с медицинской точки зрения в современном человеческом обществе.Наиболее известным примером этого типа рассуждений является гипотеза «бережливых генов», 7 , которая постулирует, что полиморфизмы, которые предрасполагают современных людей к ожирению и в настоящее время вредны с медицинской точки зрения, могут достигать высокой частоты в популяции из-за связанной избирательной преимущества во времена скудных источников пищи. Еще один механизм, уравновешивающий отбор, также может поддерживать вредные мутации в популяции, если гетерозиготные особи имеют сильное эволюционное преимущество. 8 Классическим примером этого механизма является мутация гемоглобина, защищающая от малярии в гетерозиготном состоянии и одновременно приводящая к серповидноклеточной анемии в гомозиготном состоянии.
6 Человеческий образ жизни, окружающая среда и питание резко изменились, и некоторые мутации, которые были нейтральными или даже полезными в обществах охотников-собирателей десятки тысяч лет назад, могут стать вредными с медицинской точки зрения в современном человеческом обществе.Наиболее известным примером этого типа рассуждений является гипотеза «бережливых генов», 7 , которая постулирует, что полиморфизмы, которые предрасполагают современных людей к ожирению и в настоящее время вредны с медицинской точки зрения, могут достигать высокой частоты в популяции из-за связанной избирательной преимущества во времена скудных источников пищи. Еще один механизм, уравновешивающий отбор, также может поддерживать вредные мутации в популяции, если гетерозиготные особи имеют сильное эволюционное преимущество. 8 Классическим примером этого механизма является мутация гемоглобина, защищающая от малярии в гетерозиготном состоянии и одновременно приводящая к серповидноклеточной анемии в гомозиготном состоянии.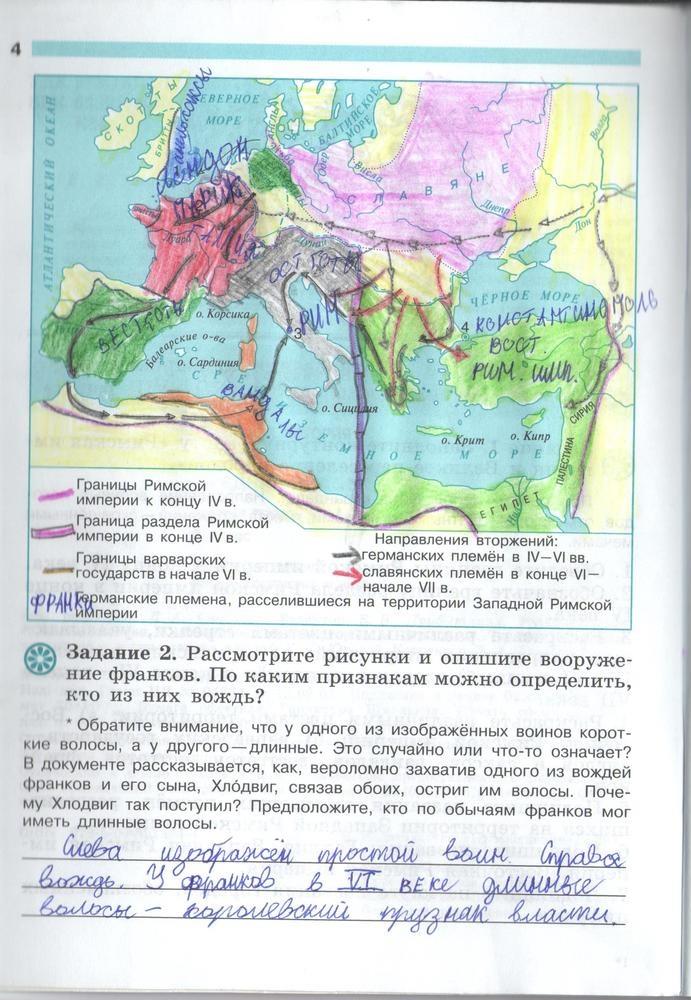 9 Четвертым явлением, которое может привести к высокой частоте вредных для здоровья мутаций, является антагонистическая плейотропия, при которой отрицательное влияние мутации на один признак компенсируется ее положительным влиянием на другой. 10 Однако ни один из этих эволюционных сценариев не оказался достаточно частым, чтобы объяснить большое количество распространенных сложных заболеваний человека.
9 Четвертым явлением, которое может привести к высокой частоте вредных для здоровья мутаций, является антагонистическая плейотропия, при которой отрицательное влияние мутации на один признак компенсируется ее положительным влиянием на другой. 10 Однако ни один из этих эволюционных сценариев не оказался достаточно частым, чтобы объяснить большое количество распространенных сложных заболеваний человека.
Вторая теория постулирует, что большинство вредных для здоровья полиморфизмов также слегка вредны для эволюции, а наблюдаемая частота предрасполагающих к болезни генетических вариаций является результатом баланса мутаций и отбора. Ключевым предположением здесь является то, что большинство болезнетворных мутаций вредны как с медицинской точки зрения, так и с точки зрения эволюции, но давление очищающего отбора, действующего на них, достаточно слабое. Высокий уровень умеренно вредных мутаций, связанных с риском заболевания, уравновешивает действие очищающего отбора.
Баланс между мутациями и отбором изначально рассматривался Кимурой теоретически. 11 Затем его защищали как 12 , так и возражали против 13 как механизма, способного поддерживать высокий уровень генетической изменчивости в природных популяциях. В последнее десятилетие, отчасти из-за популярности гипотезы «общее заболевание — общий вариант», баланс мутаций и отбора часто игнорировался как механизм, который может объяснить существование многих распространенных, вредных, наследственных заболеваний.Однако недавно баланс мутаций и отбора снова был выдвинут в качестве правдоподобного механизма поддержания вредоносной генетической изменчивости. 14 – 16
Очевидно, что баланс мутаций и отбора может стать возможным эволюционным объяснением обычных болезней только в том случае, если достаточная часть мутаций de novo у людей носит умеренно вредный характер. Предметом данной работы является оценка доли умеренно вредных миссенс-мутаций и соответствующей доли среди низкочастотных полиморфизмов человека.
Эволюционное происхождение современного пагубного полиморфизма имеет важное значение для спектра аллелей, предрасполагающих к заболеванию. Если бы большинство вредных для здоровья мутаций не были вредными с точки зрения эволюции, они могли бы получить высокую частоту в популяции, и, таким образом, гипотеза общего заболевания — общего варианта жизнеспособна. Напротив, если большинство вредных для здоровья полиморфизмов являются умеренно вредными, а их кумулятивная высокая частота в популяции поддерживается за счет баланса мутаций и отбора, то альтернативная гипотеза распространенного заболевания и редкого варианта, вероятно, будет верной.Учитывая высокий уровень заболеваемости многими сложными заболеваниями, гипотеза баланса мутаций и отбора может быть реализована только в том случае, если большая часть мутаций de novo связана с умеренными коэффициентами отбора, так что, несмотря на то, что они вредны, они все же могут достигать обнаруживаемой частоты в человеческое население.
Предыдущие теоретические исследования в рамках мутационно-селекционной модели распространенных заболеваний были сосредоточены на частотном спектре аллелей восприимчивости.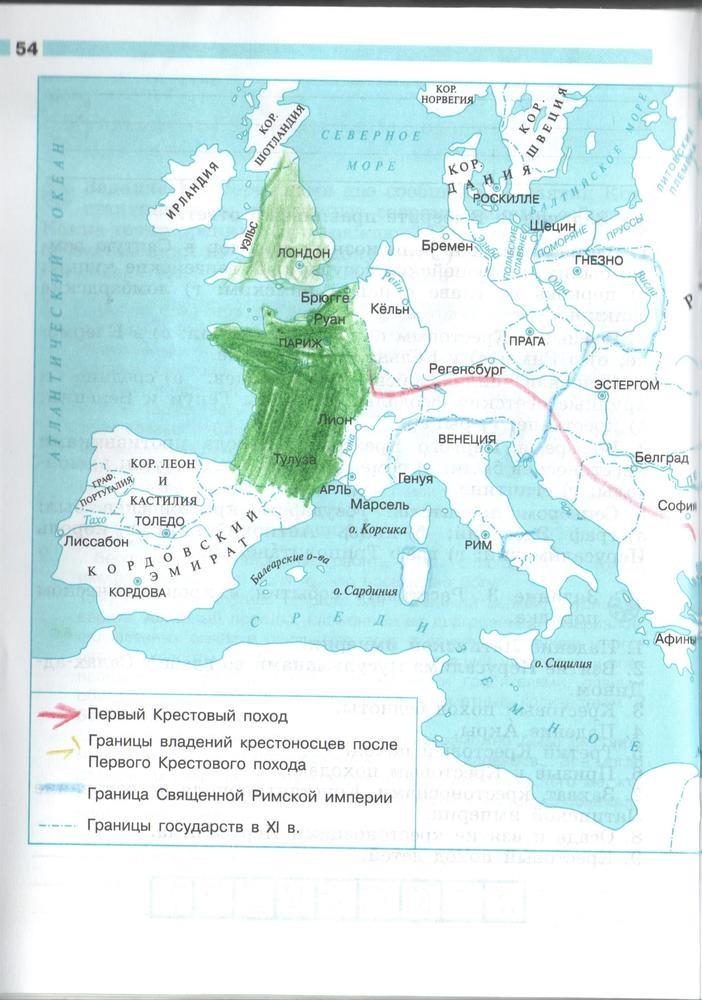 Pritchard 17 и Pritchard and Cox 18 утверждали, что низкочастотные аллели вносят основной вклад в общее заболевание, тогда как Reich и Lander 19 отстаивали гипотезу общего заболевания – распространенного варианта.Pritchard 17 предположил, что частота мутаций восприимчивости высока и что они находятся под давлением слабого очищающего отбора, что приводит к обилию редких вариантов. Райх и Ландер 19 не заостряли внимание на вопросе о том, как отдельный вредоносный вариант может достигать высокой частоты в популяции. Учитывая высокую частоту фенотипа, они рассмотрели аллельную идентичность в классе болезни, которая зависела от скорости мутаций, придающих предрасположенность к заболеванию.Таким образом, обе модели зависят от скорости вредных мутаций, связанных с болезнью, и от силы отбора против новых мутаций. В обоих исследованиях использовались оценки частоты мутаций, основанные на примерах из нескольких локусов и мутаций, которые вызывают полностью пенетрантные менделевские фенотипы; мутации, связанные с комплексным заболеванием, с другой стороны, могут иметь меньшие последствия.
Pritchard 17 и Pritchard and Cox 18 утверждали, что низкочастотные аллели вносят основной вклад в общее заболевание, тогда как Reich и Lander 19 отстаивали гипотезу общего заболевания – распространенного варианта.Pritchard 17 предположил, что частота мутаций восприимчивости высока и что они находятся под давлением слабого очищающего отбора, что приводит к обилию редких вариантов. Райх и Ландер 19 не заостряли внимание на вопросе о том, как отдельный вредоносный вариант может достигать высокой частоты в популяции. Учитывая высокую частоту фенотипа, они рассмотрели аллельную идентичность в классе болезни, которая зависела от скорости мутаций, придающих предрасположенность к заболеванию.Таким образом, обе модели зависят от скорости вредных мутаций, связанных с болезнью, и от силы отбора против новых мутаций. В обоих исследованиях использовались оценки частоты мутаций, основанные на примерах из нескольких локусов и мутаций, которые вызывают полностью пенетрантные менделевские фенотипы; мутации, связанные с комплексным заболеванием, с другой стороны, могут иметь меньшие последствия. Различие в выводах объясняется главным образом различием в численных значениях скорости вредных мутаций и силы действующего на них отбора.Следует подчеркнуть, что частота вредоносных мутаций зависит не только от исходной скорости замещения каждого нуклеотида, но также и от доли вредоносных мутаций de novo. Надежная оценка этих параметров требует интеграции нескольких типов данных.
Различие в выводах объясняется главным образом различием в численных значениях скорости вредных мутаций и силы действующего на них отбора.Следует подчеркнуть, что частота вредоносных мутаций зависит не только от исходной скорости замещения каждого нуклеотида, но также и от доли вредоносных мутаций de novo. Надежная оценка этих параметров требует интеграции нескольких типов данных.
В нашем анализе мы объединили данные о мутациях, вызывающих менделевскую болезнь человека, дивергенции человека и шимпанзе и генетической изменчивости в современной человеческой популяции. Наличие новых данных о полиморфизме человека, обнаруженном в 37 связанных с ожирением генах, секвенированных у 756 человек, позволило нам исследовать важный класс умеренно вредных мутаций.Очищающий отбор, действующий на этот тип мутаций, достаточно силен, чтобы эффективно предотвратить их фиксацию, но они присутствуют в популяции с низкой частотой и могут быть обнаружены при секвенировании большого числа особей.
Мы подсчитали, что >50% миссенс-мутаций de novo в среднем гене человека и 70% миссенс-SNP, обнаруженных только один раз среди 1500 хромосом, являются умеренно вредными. Такие умеренно вредные мутации связаны с коэффициентами отбора в удивительно узком диапазоне от 0.001–0,003.
Такие умеренно вредные мутации связаны с коэффициентами отбора в удивительно узком диапазоне от 0.001–0,003.
Высокая доля умеренно вредных мутаций среди миссенс-мутаций предполагает, что баланс мутации-отбора может быть возможным объяснением существования распространенного заболевания со сложным наследованием. Наблюдение, что большинство редких несинонимичных вариантов человека являются вредными и, таким образом, имеют значение для функции и фенотипа, сильно поддерживает стратегию повторного секвенирования для исследований ассоциации генов-кандидатов; Ожидается, что в больной популяции будет более высокий уровень редких вариантов аминокислот в генах, участвующих в заболевании, чем в здоровой контрольной группе.
. Данные и методы. миссенс, нонсенс и мутации сайтов сплайсинга. Связанные с заболеванием полиморфизмы составляют лишь очень небольшую часть HGMD, и большинство мутаций, включенных в базу данных, являются полностью пенетрантными и вызывают простые менделевские заболевания.
 20 В HGMD отсутствует информация о том, сколько раз была идентифицирована каждая отдельная мутация. Однако, если предположить, что мутации следуют статистике Пуассона, наличие мутаций, обнаруженных несколько раз, не окажет существенного влияния на наши оценки, если только не будет обнаружено большинство всех возможных бессмысленных мутаций, что крайне маловероятно для большинства генов.
20 В HGMD отсутствует информация о том, сколько раз была идентифицирована каждая отдельная мутация. Однако, если предположить, что мутации следуют статистике Пуассона, наличие мутаций, обнаруженных несколько раз, не окажет существенного влияния на наши оценки, если только не будет обнаружено большинство всех возможных бессмысленных мутаций, что крайне маловероятно для большинства генов.
HGMD может включать гены с полной потерей функции мутаций, которые являются летальными для эмбриона. Он содержит некоторые мутации с приобретением функции и мутации с неполной пенетрантностью.Хотя доля этих мутаций в HGMD, вероятно, невелика, их наличие может привести к смещению наших оценок. Таким образом, мы повторили наш анализ на меньшем, но хорошо охарактеризованном наборе генов аутосомно-доминантных или Х-сцепленных заболеваний, первоначально собранных Kondrashov 21 из локус-специфических баз данных для оценки частоты мутаций у людей. Мы извлекли информацию о миссенс- и нонсенс-мутациях в этих генах из отдельных записей HGMD. Чтобы гарантировать, что полная потеря функции не является летальной, мы ограничили наш анализ 26 генами, которые имели по крайней мере пять зарегистрированных бессмысленных мутаций.Считается, что все миссенс-мутации в этих генах являются мутациями с потерей функции.
Чтобы гарантировать, что полная потеря функции не является летальной, мы ограничили наш анализ 26 генами, которые имели по крайней мере пять зарегистрированных бессмысленных мутаций.Считается, что все миссенс-мутации в этих генах являются мутациями с потерей функции.
Данные о полиморфизме человека: новый большой набор данных повторного секвенирования
Для нашего анализа очень низкочастотных несинонимичных SNP мы использовали новый большой набор данных повторного секвенирования, описанный Ahituv et al. (в этом выпуске). 22 Полные экзонические последовательности и их сайты сплайсинга были секвенированы в 58 генах с потенциальным участием в ожирении у 379 тучных и 378 худых людей. Полное секвенирование >1500 хромосом дало нам возможность изучить несинонимичные SNP с очень низкой частотой.Все лица, включенные в исследование, имеют белое происхождение.
Поскольку этот набор данных не основан на случайной популяционной выборке и имеет фенотипическую предвзятость, мы ограничили наш анализ 37 аутосомными генами, для которых не было обнаружено доказательств влияния на ожирение. В этих генах не было статистически значимого превышения вариации ни в группе с ожирением, ни в группе с худощавым телом; 71 и 79 миссенс-вариантов были обнаружены в худых и тучных когортах соответственно. Случайная повторная выборка подмножеств, соответствующих набору данных SeattleSNP по размеру, дала оценки, очень похожие на оценки, полученные из меньших систематических наборов данных, что еще раз подтвердило отсутствие систематической ошибки в выбранном подмножестве из 37 генов.
В этих генах не было статистически значимого превышения вариации ни в группе с ожирением, ни в группе с худощавым телом; 71 и 79 миссенс-вариантов были обнаружены в худых и тучных когортах соответственно. Случайная повторная выборка подмножеств, соответствующих набору данных SeattleSNP по размеру, дала оценки, очень похожие на оценки, полученные из меньших систематических наборов данных, что еще раз подтвердило отсутствие систематической ошибки в выбранном подмножестве из 37 генов.
Данные о полиморфизме человека: общедоступные данные
В дополнение к нашему новому набору данных повторного секвенирования мы использовали три общедоступных набора данных, которые содержат достаточную информацию о редких несинонимичных аллелях: (1) набор данных, созданный Национальным институтом гигиены окружающей среды. Science Environmental Genome Project (NIEHS-EGP) (веб-сайт программы NIEHS SNPs), 23 , в котором >500 генов, участвующих в репарации ДНК и путях клеточного цикла, были секвенированы по меньшей мере у 90 неродственных людей; (2) набор данных, созданный проектом SeattleSNPs (SeattleSNPs), в котором >200 генов, участвующих в воспалительных реакциях, были секвенированы по крайней мере у 46 человек; и (3) набор данных, созданный базой данных японских одиночных нуклеотидных полиморфизмов ( JSNP ), 24 , в которой полиморфные сайты в более чем 8000 генах были обнаружены с использованием панели из 12 человек, а затем были генотипированы у 750 человек.
Мутации, зафиксированные в человеческой линии после дивергенции от шимпанзе
Для получения информации о мутациях, зафиксированных в человеческой линии после дивергенции от шимпанзе, мы использовали парные выравнивания полных геномов человека-шимпанзе и человека-макаки, построенные с помощью программы BLASTZ. Эти выравнивания были получены с сайта Калифорнийского университета в Санта-Круз (UCSC) (UCSC Genome Bioinformatics: Sequence and Annotation Downloads). Мы сравнили все нуклеотидные замены, обнаруженные между полными геномами человека и шимпанзе, с базой данных dbSNP , 25 , которая содержит информацию о большинстве известных генетических вариаций человека.Замены, которые были обнаружены как современные полиморфизмы, были исключены из нашего анализа дивергенции человека и шимпанзе. Последовательность макак использовалась в качестве внешней группы, чтобы отличить мутации, зафиксированные в линии человека, от мутаций, зафиксированных в линии шимпанзе. Соответственно, мы игнорировали сайты, в которых нуклеотид макака отличался от нуклеотидов человека и шимпанзе. Из-за этой процедуры мы потенциально исключили небольшую часть очень быстро развивающихся сайтов, тем самым слегка недооценив долю «эффективно нейтральных» аминокислотных замен.
Соответственно, мы игнорировали сайты, в которых нуклеотид макака отличался от нуклеотидов человека и шимпанзе. Из-за этой процедуры мы потенциально исключили небольшую часть очень быстро развивающихся сайтов, тем самым слегка недооценив долю «эффективно нейтральных» аминокислотных замен.
Контекстно-зависимая модель мутаций
Для точной оценки селективных ограничений в областях, кодирующих белок, требуется контекстно-зависимая мутационная модель. 26 Мы построили контекстно-зависимую матрицу частоты мутаций, которая описывала вероятность возникновения мутации de novo до того, как имел место какой-либо отбор. С помощью такой модели мутабельности мы смогли сравнить наблюдаемое количество различных замен с соответствующим числом, ожидаемым при нейтральности, и из такого сравнения сделать вывод о силе очищающего отбора.
Мы ограничили наш анализ однонуклеотидными заменами, исключив другие менее распространенные типы мутаций. Реальный мутационный спектр довольно сложен — CpG-динуклеотиды имеют скорость мутаций на порядок выше, чем у других динуклеотидов; переходы заметно чаще, чем трансверсии; Сообщалось, что существуют некоторые другие, более тонкие, зависящие от контекста эффекты.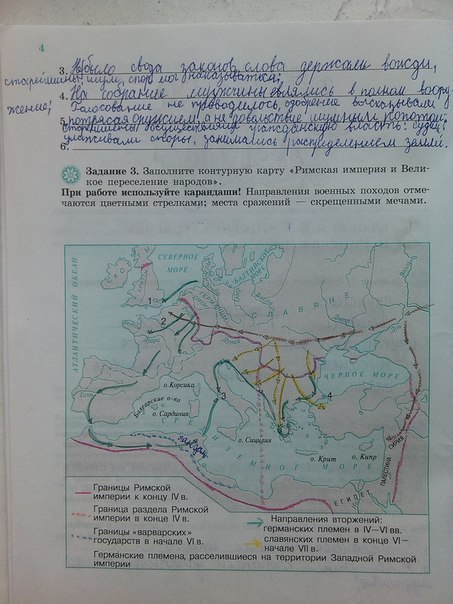 27 В других местах использовались различные модели мутаций разной степени сложности. 28 Доступность геномных последовательностей человека, шимпанзе и (неполного) бабуина позволила нам рассчитать эмпирическую «направленную» матрицу мутаций 64×3 для триплетов (с вероятностями для всех XY 1 Z до XY 2 Z одиночных -нуклеотидные замены). Такая мутационная модель, учитывающая нуклеотид, его ближайших соседей и направление замещения, должна отражать большинство известных контекстных зависимостей мелкомасштабной скорости мутаций. Мы использовали модель зависимости второго порядка от соседних позиций, чтобы зафиксировать все зависящие от контекста эффекты на частоту мутаций.Мы также ввели статистическую поправку на обратные замены в линии бабуинов.
27 В других местах использовались различные модели мутаций разной степени сложности. 28 Доступность геномных последовательностей человека, шимпанзе и (неполного) бабуина позволила нам рассчитать эмпирическую «направленную» матрицу мутаций 64×3 для триплетов (с вероятностями для всех XY 1 Z до XY 2 Z одиночных -нуклеотидные замены). Такая мутационная модель, учитывающая нуклеотид, его ближайших соседей и направление замещения, должна отражать большинство известных контекстных зависимостей мелкомасштабной скорости мутаций. Мы использовали модель зависимости второго порядка от соседних позиций, чтобы зафиксировать все зависящие от контекста эффекты на частоту мутаций.Мы также ввели статистическую поправку на обратные замены в линии бабуинов.
Мы провели выравнивание последовательностей человека, шимпанзе и бабуина, взятых из областей ENCODE, 29 , с использованием выравнивателя множественных последовательностей TBA. 30 Были подсчитаны частоты всех триплетов нуклеотидов в выровненных последовательностях (любые триплеты, содержащие пробелы, были проигнорированы). Направленность мутации (независимо от того, произошла ли замена в линии человека или шимпанзе) определяли с помощью последовательности бабуина.
Направленность мутации (независимо от того, произошла ли замена в линии человека или шимпанзе) определяли с помощью последовательности бабуина.
Вычисленная матрица согласуется с другими недавно опубликованными контекстно-зависимыми моделями нейтральной эволюции. 31 – 33 Например, коэффициент корреляции r 2 между нашей матрицей и матрицей Зипеля и Хаусслера 31 составляет 96%.
Наборы генов
Популяционная и эволюционная динамика генов, расположенных на половых хромосомах, отличается от остального генома. Чтобы избежать ненужных осложнений в нашем анализе, мы использовали только аутосомные гены.Для расчета соотношения несинонимичных и синонимичных мутаций, характерных для всего генома человека, мы использовали 14 095 надежно аннотированных аутосомных генов из проекта Consensus CDS (CCDS).
Достоверность оценок
Анализ дивергенции человека и шимпанзе основан на полном протеоме, а анализ базы данных HGMD включает наиболее полный набор генов болезней. Однако данные SNP человека представлены гораздо меньшими наборами генов. Следовательно, необходимо присвоить достоверность статистическим оценкам, полученным из этих наборов.Несмотря на то, что эти наборы данных содержат большое количество SNP, мутации были обнаружены в разных генах, обладающих разными свойствами. Таким образом, ожидается, что ошибки выборки оценок будут в основном определяться выборкой генов, а не отдельных мутаций.
Однако данные SNP человека представлены гораздо меньшими наборами генов. Следовательно, необходимо присвоить достоверность статистическим оценкам, полученным из этих наборов.Несмотря на то, что эти наборы данных содержат большое количество SNP, мутации были обнаружены в разных генах, обладающих разными свойствами. Таким образом, ожидается, что ошибки выборки оценок будут в основном определяться выборкой генов, а не отдельных мутаций.
Мы вычислили SE из SD оценок, полученных из случайных непересекающихся подмножеств данных. Хотя SE в основном используются для оценки среднего, мы проверили в серии симуляций, что SE, вычисленные с использованием непересекающихся подмножеств, удовлетворительно аппроксимируют ошибку оценок, представленных в этой работе.
Результаты
Сильно вредные мутации
В качестве первого шага в нашем анализе спектра потенциальных эффектов аминокислотных мутаций мы оценили долю миссенс-изменений de novo, которые являются сильно вредными. Мы определяем миссенс-мутацию как сильно вредную, если она вызывает полную потерю функции белка, что часто наблюдается при менделевских заболеваниях.
Мы определяем миссенс-мутацию как сильно вредную, если она вызывает полную потерю функции белка, что часто наблюдается при менделевских заболеваниях.
Бессмысленные мутации, вводящие преждевременные стоп-кодоны, могут служить показателем «сильного вреда».Всего в HGMD указано 26 305 миссенс-мутаций и 6764 нонсенс-мутации. Хотя HGMD имеет врожденную предвзятость установления, следуя работе Kondrashov 21 и Yampolsky et al., 34 , мы считаем, что мутации, вызывающие один и тот же фенотип, с одинаковой вероятностью будут помещены в базу данных, независимо от их типа — ерунда, миссенс, или сайт сплайсинга. Если бы все миссенс-мутации с такой же вероятностью приводили к полной потере функции и впоследствии к выраженному фенотипу заболевания, как и нонсенс-мутации, то отношение миссенс-мутаций к нонсенс-мутациям в базе данных HGMD было бы подобно ожидаемому теоретическому соотношению для de novo. мутации.В реальности она значительно ниже. Используя нашу модель частоты мутаций, описанную выше, мы подсчитали, что ∼19,7 миссенс-замен происходят в масштабе всего генома на каждую нонсенс-замену (для расчета полногеномных значений было использовано 14 095 генов человека из проекта CCDS ). В то же время среди болезнетворных мутаций в базе данных HGMD на одну нонсенс-замену приходилось всего 3,9 миссенс-изменения (). Такое различие указывает на то, что большая часть миссенс-мутаций недостаточно опасна, чтобы быть видимой через такой «сильный фенотипический фильтр», как обнаружение и включение в HGMD.Используя полученные числа, мы можем оценить, что 20% (3,9 из 19,7) миссенс-мутаций являются сильно вредными.
В то же время среди болезнетворных мутаций в базе данных HGMD на одну нонсенс-замену приходилось всего 3,9 миссенс-изменения (). Такое различие указывает на то, что большая часть миссенс-мутаций недостаточно опасна, чтобы быть видимой через такой «сильный фенотипический фильтр», как обнаружение и включение в HGMD.Используя полученные числа, мы можем оценить, что 20% (3,9 из 19,7) миссенс-мутаций являются сильно вредными.
Спектр эффектов миссенс-мутаций de novo. A, Доля сильно вредных мутаций среди аминокислотных замен de novo. Нонсенс-мутации в HGMD, вызывающие заболевание, использовались в качестве стандарта «сильного вреда». B, Доля сильно вредных мутаций среди аминокислотных замен de novo. Болезнетворные мутации сайта сплайсинга в HGMD использовались в качестве стандарта «сильного вреда».” C, Доля эффективно нейтральных мутаций среди аминокислотных замен de novo. Синонимические замены, зафиксированные в человеческой линии после дивергенции от шимпанзе, использовались в качестве эталона «эффективной нейтральности».
Мутации, которые нарушают динуклеотиды основного сайта сплайсинга (GT/AG), часто приводят к почти полной потере функции гена. Эти замены сайта сплайсинга можно использовать в качестве «эталона» для сильно вредных мутаций так же, как мы использовали бессмысленные мутации.Мы подсчитали, что по всему геному ожидается ~36,5 миссенс-мутаций de novo на одну мутацию сайта сплайсинга de novo. Однако в HGMD только 7,6 миссенс-мутаций перечислены на мутацию в сайте сплайсинга (1). Соответственно, мы можем оценить, что доля сильно вредных мутаций среди миссенс-мутаций составляет ∼21%. Это число находится в замечательном соответствии со значением, полученным с использованием бессмысленных мутаций в качестве эталона.
Хотя HGMD обеспечивает исчерпывающий набор мутаций заболевания, он очень гетерогенен и включает мутации неполной пенетрантности, мутации с приобретением функции и, возможно, мутации в генах с полной потерей функции для эмбриона.Наличие мутаций в двух последних категориях сместило бы нашу оценку в сторону увеличения, то есть привело бы к консервативной оценке для целей данной работы. Однако, чтобы проверить, может ли быть значительным смещение вверх, мы проанализировали гораздо меньший набор очень хорошо охарактеризованных генов, участвующих в аутосомно-доминантных или Х-сцепленных простых менделевских заболеваниях. Считается, что все мутации в этом наборе приводят к потере функции, и для них полная потеря функции не смертельна. После того, как были приняты во внимание длины отдельных генов, 21% новых миссенс-мутаций в наборе были оценены как сильно вредные.
Однако, чтобы проверить, может ли быть значительным смещение вверх, мы проанализировали гораздо меньший набор очень хорошо охарактеризованных генов, участвующих в аутосомно-доминантных или Х-сцепленных простых менделевских заболеваниях. Считается, что все мутации в этом наборе приводят к потере функции, и для них полная потеря функции не смертельна. После того, как были приняты во внимание длины отдельных генов, 21% новых миссенс-мутаций в наборе были оценены как сильно вредные.
Эффективно нейтральные мутации
В качестве второго шага в нашем анализе спектра потенциальных эффектов аминокислотных замен мы оценили долю миссенс-мутаций de novo, которые являются эффективно нейтральными. Мы определяем миссенс-мутацию как эффективно нейтральную, если вероятность ее закрепления в предковой человеческой популяции после дивергенции от шимпанзе была бы аналогична вероятности синонимичных замен. Хотя было показано, что синонимические замены не являются полностью избирательно нейтральными 35 – 37 (обзор Chamary et al. 38 ), считается, что влияние на приспособленность подавляющего большинства из них в человеческой популяции относительно невелико.
38 ), считается, что влияние на приспособленность подавляющего большинства из них в человеческой популяции относительно невелико.
По нашим оценкам (), на одну синонимическую мутацию приходится 2,23 миссенс-мутации de novo. Однако среди замен, зафиксированных в линии человека после дивергенции от шимпанзе (рассчитанной с использованием геномной последовательности макаки в качестве внешней группы), на одну синонимическую замену приходилось только 0,60 миссенс-мутаций. Остальные миссенс-мутации, по-видимому, были устранены очищающим отбором.Исходя из этих значений, мы можем оценить, что ∼27% (0,6 из 2,23) миссенс-мутаций в белках человека аналогичны по действию синонимичным заменам.
Эта оценка чувствительна к полезным мутациям, закрепившимся за счет положительного отбора, и к очень слабо вредным мутациям, закрепившимся в результате дрейфа. Наличие этих мутаций сместило бы нашу оценку доли умеренно вредных мутаций в сторону понижения, то есть наша оценка является консервативной для наших целей. Он также консервативен в отношении очищающего отбора по синонимичным сайтам.Таким образом, эту оценку можно рассматривать как оценку доли новых миссенс-мутаций, которые не связаны с потерей приспособленности, большей, чем величина, обратная эффективному размеру популяции.
Он также консервативен в отношении очищающего отбора по синонимичным сайтам.Таким образом, эту оценку можно рассматривать как оценку доли новых миссенс-мутаций, которые не связаны с потерей приспособленности, большей, чем величина, обратная эффективному размеру популяции.
Слабо вредные полиморфизмы
Мы проанализировали два крайних значения потенциального воздействия изменений аминокислот на приспособленность и подсчитали, что среди миссенс-мутаций de novo в белках человека примерно 20% являются сильно вредными, тогда как другие 27% являются практически нейтральными. Простая арифметика приводит нас к выводу, что большинство (53%) всех мутаций de novo missense на самом деле умеренно вредны.Мягко вредные мутации могут достигать низких, но поддающихся обнаружению частот в популяции. Затем мы задали вопрос: если доля умеренно вредных мутаций среди мутаций de novo составляет 53%, какова доля умеренно вредных аллелей среди редких и распространенных несинонимичных SNP в человеческой популяции?
Во-первых, мы рассмотрели очень редкие миссенс-полиморфизмы. Мы проанализировали только те полиморфные сайты, в которых успешно секвенировано не менее 1400 из 1514 хромосом. Более 60% несинонимичных SNP, обнаруженных в нашем наборе из 37 аутосомных генов, секвенированных у 757 человек, были обнаружены как одиночные (т.э., были обнаружены в гетерозиготном состоянии у одной особи). Несмотря на то, что синглетоны представляют собой SNP с очень низкой частотой, вероятность того, что они являются мутациями de novo, становится все меньше. Количество несинонимичных SNP, выявляемых с более высокими частотами, было значительно меньше, и, таким образом, синглетоны представляли собой единственную группу редких миссенс-аллелей, по которой было достаточно данных для статистически значимого анализа.
Мы проанализировали только те полиморфные сайты, в которых успешно секвенировано не менее 1400 из 1514 хромосом. Более 60% несинонимичных SNP, обнаруженных в нашем наборе из 37 аутосомных генов, секвенированных у 757 человек, были обнаружены как одиночные (т.э., были обнаружены в гетерозиготном состоянии у одной особи). Несмотря на то, что синглетоны представляют собой SNP с очень низкой частотой, вероятность того, что они являются мутациями de novo, становится все меньше. Количество несинонимичных SNP, выявляемых с более высокими частотами, было значительно меньше, и, таким образом, синглетоны представляли собой единственную группу редких миссенс-аллелей, по которой было достаточно данных для статистически значимого анализа.
Мы оценили относительную долю миссенс-мутаций de novo, представленных одиночками.Эта оценка была основана на сравнении наблюдаемого числа несинонимичных замен на синонимическую мутацию (отношение N a / N s ) с соответствующим теоретическим числом, ожидаемым при нейтральной эволюции ( N ). 0 a / N 0 s соотношение). Следует отметить, что предположение об отсутствии естественного отбора (нейтральная эволюция) равносильно предположению, что естественный отбор еще не действовал (мутации de novo).
0 a / N 0 s соотношение). Следует отметить, что предположение об отсутствии естественного отбора (нейтральная эволюция) равносильно предположению, что естественный отбор еще не действовал (мутации de novo).
, как представлено и,
3 N 0 0 / N / N / N 0 0 S Соотношение, рассчитанное с использованием нашей нейтральной модели эволюции для 37 генов большого контрольного набора данных, было равно 2.204. Экспериментальное отношение N a / N s для синглетонов, рассчитанное для того же набора данных, равно 1,49. Учитывая эти значения, мы определили, что только 32% (1-1.49/2.20) миссенс-мутаций вредны в той мере, в какой существует очень низкая вероятность того, что они будут обнаружены хотя бы один раз в образце из 1500 хромосом. SE этой оценки составляет 2%.
SE этой оценки составляет 2%.
Доля миссенс-мутаций de novo, представленных на разных уровнях частоты аллелей. Нормализованная доля аминокислотных замен de novo, обнаруженных в заданном наборе данных, была рассчитана по разнице наблюдаемого соотношения N a / N s и теоретического N 7 a / N 0 s отношение, ожидаемое при нейтральности.Данные по редким полиморфизмам показаны оранжевым цветом, данные по распространенным полиморфизмам — желтым, а данные по заменам (субст.), зафиксированным в человеческой линии после расхождения с шимпанзе, — зеленым. SE показаны серыми полосами погрешностей.
Таблица 1.
Соотношения распространения 3
16
|
16 |
6 |
17 | N N / N / S S ( N / N S ) / ( N 0 A / N 0 S )
|
||||||||||||||||||||||||||||||
| Набор данных | Описание набора данных | №Генов | Теоретический N 0 0 / N / N 0 0 S S 8 Singletons |
Snps с MAF> 25% |
Замена в Линии человека |
Singletons |
SNP с MAF> 25% A |
Замена в Lineage A A |
| ||||||||||||||||||||||||
| CCDS | Genomewide | 14 095 | 2. |
… | … | … | .60 | . | .44 | .68 ± 0,0424 .68 ± 0,02244. 68 ± 0,02 | … | … | .20 ± 0,0516 | Niehs-EGP | 90-95 отдельных лиц секвенировали | 518 | 1. 255 255 |
. 56 | .50 | .61 ± 0,0424 .61 ± 0,02424 .61 ± 0,02 | .26 ± 0,04224,26 ± 0,04 | . 22 ± 0,0116 | STEATLESNPS | 46-47 человек секвенировали | 236 | 236 | 2.203 | 1.43 | .66 | . 66 | . 65 ± 0,0424 0,65 ± 0,04 65 ± 0,0424 0,65 ± 0,04 |
.32 ± 0,05 | . 30318 |
| jsnp | 750 человек генотипирован | 8,786 | 2.244 | 1.244 | 1,37 . 5440318
| .54 |
.54 |
.61 |
.24 |
.24 |
. |
 24 24 .24 |
|
Экспериментальный N A / N / N / N / S / S Соотношение Соотношение аминокислотных замещений, закрепленных в наборе 37 генов в людской линии после расхождения от шимпанзе равно 0,44. Это означает, что примерно 20% всех миссенс-мутаций в этих генах эффективно нейтральны — значение, близкое к среднему по геному.
Генетическая изменчивость человека, обнаруженная в наборе из 37 генов, секвенированных у 756 человек, предоставила возможность получить представление о генетической изменчивости человека с очень низкой частотой аллелей.Однако ему не хватает значительного количества данных об общей генетической изменчивости. Чтобы восполнить этот пробел, мы использовали три общедоступных набора данных: (1) набор данных, созданный NIEHS-EGP, в котором > 500 генов, участвующих в репарации ДНК и путях клеточного цикла, были секвенированы по крайней мере у 90 неродственных людей; (2) набор данных, созданный SeattleSNP, в котором > 200 генов, участвующих в воспалительных реакциях, были секвенированы по крайней мере у 46 человек; и (3) набор данных, созданный JSNP, в котором полиморфные сайты в более чем 8000 генах были обнаружены с использованием панели из 12 человек, а затем были генотипированы у 750 человек. Эти наборы данных также содержали информацию о редкой генетической изменчивости, которую мы использовали, хотя большая часть редких несинонимичных SNP с частотой <1% могла быть пропущена в наборах данных NIEHS-EGP и SeattleSNP, поскольку было секвенировано <200 хромосом и, возможно, пропущены в JSNP, потому что для обнаружения SNP использовались только 12 человек.
Эти наборы данных также содержали информацию о редкой генетической изменчивости, которую мы использовали, хотя большая часть редких несинонимичных SNP с частотой <1% могла быть пропущена в наборах данных NIEHS-EGP и SeattleSNP, поскольку было секвенировано <200 хромосом и, возможно, пропущены в JSNP, потому что для обнаружения SNP использовались только 12 человек.
Снова, используя нашу нейтральную модель эволюции, мы рассчитывали предсказанные N 0 N / N / N 0 0 S Соотношение отдельно для Niehs-EGP, STECTLESNPS и JSNP Наборы данных .Затем мы рассчитали экспериментальные отношения N a / N s для трех типов смены последовательностей. Первый тип — это редкие несинонимичные SNP, которые либо ( a ) наблюдались только один раз среди всех секвенированных хромосом (в NIEHS-EGP и Seattle SNP), либо ( b ) имели наблюдаемую частоту <1% (JSNP).
Мы наблюдали, что N / N N N S S Соотношения для очень распространенных полиморфизмов были только очень чуть выше N A / N S соотношения для фиксированных замен ( и ). Этот факт указывает на то, что доля вредных аминокислотных замен среди обычных SNP очень мала. Это также подтверждает мнение о том, что доля положительно выбранных аминокислотных замен среди всех мутаций в белках человека была низкой. 39
в резком контрасте с обыкновенными SNP, N A N / N / N Соотношение на очень редкие аллели значительно превышает N A / N N N N s коэффициенты для фиксированных замен (предположительно нейтральных). Этот факт указывает на то, что очень большая часть таких редких миссенс-SNP является вредной. Фракция вредных мутаций среди наблюдаемых редких замеек может быть оценена как ( N Red A / N Red S ) / ( N a / N фиксированный s ).Простые расчеты показывают, что большинство (52–71%) аминокислотных замен с наблюдаемой частотой <1% являются умеренно вредными во всех наборах данных (1). Это неожиданное открытие указывает на то, что низкая частота миссенс-мутаций сама по себе может служить сильным предиктором вредного эффекта полиморфных вариантов.
Этот факт указывает на то, что очень большая часть таких редких миссенс-SNP является вредной. Фракция вредных мутаций среди наблюдаемых редких замеек может быть оценена как ( N Red A / N Red S ) / ( N a / N фиксированный s ).Простые расчеты показывают, что большинство (52–71%) аминокислотных замен с наблюдаемой частотой <1% являются умеренно вредными во всех наборах данных (1). Это неожиданное открытие указывает на то, что низкая частота миссенс-мутаций сама по себе может служить сильным предиктором вредного эффекта полиморфных вариантов.
Таблица 2.
Таблица 2.
Фракция вредных замещений между редкой миссией SNPS
| набор | Количество секвенированных лиц | Процент вредных SNP среди проведений проводников A | |
| Набор данных Гены, связанные с ожирением | 757 | 71 ± 8 | |
| 90-95 | 64 ± 1 | 64 ± 1 | |
| Seattlesnps | 46-47 | 52 ± 6 | 52 ± 6 |
Характеристики Коэффициенты отбора теории умеренно вредных мутаций
Мы пришли к выводу, что более половины вновь возникающих миссенс-мутаций являются умеренно вредными. Такие мутации отсутствуют среди распространенных полиморфизмов, но очень распространены среди замен, обнаруживаемых только один раз в ⩾100 хромосом. На основании этих наблюдений мы оценили коэффициенты отбора, связанные с такими мутациями.
Такие мутации отсутствуют среди распространенных полиморфизмов, но очень распространены среди замен, обнаруживаемых только один раз в ⩾100 хромосом. На основании этих наблюдений мы оценили коэффициенты отбора, связанные с такими мутациями.
Сначала мы рассчитали теоретическое ожидаемое число мутаций с коэффициентом отбора s , наблюдаемых только один раз в наборе из м случайно отобранных хромосом. Затем мы разделили его на соответствующее количество нейтральных мутаций, чтобы получить соотношение эффективный размер популяции, а x — частота отдельного аллеля в популяции.Количество обнаруженных мутаций было получено путем умножения теоретического числа всех аллелей, присутствующих в популяции с частотой х , на вероятность того, что один аллель с частотой х будет обнаружен как синглтон в наборе из м хромосом и затем путем интегрирования по частоте аллеля x от 0 до 1. Следует отметить, что аллель обнаруживается как синглтон в наборе из m хромосом, если присутствует либо 1, либо m -1 копия.
Аналогичным образом мы рассчитали теоретическое соотношение R MAF >0,25 ( s ) для мутаций, которые были обнаружены как полиморфизмы в наборе из m хромосом с частотой минорного аллеля (MAF) >25 %.
, где C n k — биномиальный коэффициент — число комбинаций размера k из набора с n результатом применения 90s теории формул 90s к динамике полиморфизма в популяциях. 40 Их два основных основных предположения заключаются в постоянном эффективном размере популяции и в том, что каждая новая мутация происходит в новом, ранее мономорфном сайте (модель с бесконечным числом сайтов). Коэффициент доминирования принимался равным 0,5, поэтому, если s — коэффициент отбора, связанный с гетерозиготой, то 2 s — коэффициент отбора, связанный с гомозиготой. Точные значения коэффициента доминирования не очень важны, пока они достаточно высоки, чтобы отбор мог действовать преимущественно на гетерозиготы.
Зависимость R MAF > 0.25 ( S ) и R 1/ M ( S ) (для различных значений м ) на коэффициенту выбора показан в . Мы определили, что примерно равные доли миссенс-мутаций могут наблюдаться как фиксированные замены и как общие полиморфизмы с MAF> 25% (1). Это наблюдение означает, что умеренно вредные мутации практически отсутствуют среди частых SNP и, следовательно, должны иметь значения коэффициента отбора, при которых R MAF >0.25 ( s ) очень близко к нулю. С другой стороны, большинство умеренно вредных мутаций имеют высокие относительные шансы быть обнаруженными как одиночные. Для каждого из трех наборов данных повторного секвенирования (связанных с ожирением, NIEHS-EGP и SeattleSNP) мы оценили долю умеренно вредных миссенс-мутаций de novo, наблюдаемых как одиночные, относительно нейтральных ожиданий:
Здесь числитель отражает наблюдаемое доля мутаций в классе умеренно вредоносных, обнаруживаемых как одиночные, равна доле всех мутаций, обнаруживаемых как одиночные, за вычетом доли эффективно нейтральных мутаций de novo («hum. лин.подст.” указывает на замену человеческого происхождения). Знаменатель отражает общую долю мутаций, возникающих в классе умеренно вредных мутаций. Его получают путем вычитания из 1 доли сильно вредных мутаций de novo (обозначаемых F сильно дел. ) и нейтральных мутаций. ) и R MAF >0,25 ( s ) (см. формулу [1]), рассчитанных с использованием уравнений, полученных из теории диффузии в предположении постоянной численности популяции и бесконечного числа участков.Ожидаемый сдвиг кривых R 1/ м ( с ) из-за недавнего расширения популяции показан красными стрелками. Черные стрелки иллюстрируют оценку характеристических коэффициентов отбора для класса умеренно вредных миссенс-мутаций (см. раздел «Результаты» и формулу [2]). B, R M M ( S ) и R R MAF > 0.25 ( S ) Рассчитаны прямым компьютерным моделированием молекулярной эволюции в предположении бесконечного числа.
лин.подст.” указывает на замену человеческого происхождения). Знаменатель отражает общую долю мутаций, возникающих в классе умеренно вредных мутаций. Его получают путем вычитания из 1 доли сильно вредных мутаций de novo (обозначаемых F сильно дел. ) и нейтральных мутаций. ) и R MAF >0,25 ( s ) (см. формулу [1]), рассчитанных с использованием уравнений, полученных из теории диффузии в предположении постоянной численности популяции и бесконечного числа участков.Ожидаемый сдвиг кривых R 1/ м ( с ) из-за недавнего расширения популяции показан красными стрелками. Черные стрелки иллюстрируют оценку характеристических коэффициентов отбора для класса умеренно вредных миссенс-мутаций (см. раздел «Результаты» и формулу [2]). B, R M M ( S ) и R R MAF > 0.25 ( S ) Рассчитаны прямым компьютерным моделированием молекулярной эволюции в предположении бесконечного числа.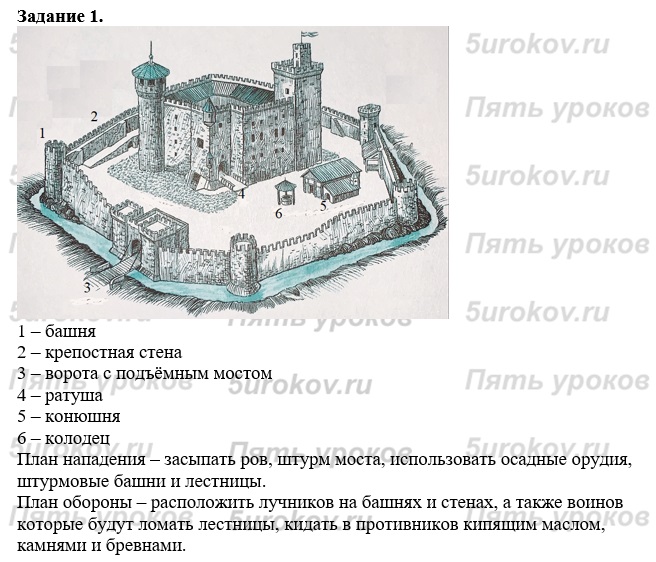 участков и простой истории населения — эпоха стабильного размера населения, за которой последовало узкое место, а затем быстрое расширение.
участков и простой истории населения — эпоха стабильного размера населения, за которой последовало узкое место, а затем быстрое расширение.
Используя теоретическую зависимость R от коэффициента отбора () для соответствующего числа секвенированных хромосом m, , мы оценили характеристические коэффициенты отбора для умеренно вредной мутации de novo миссенс для генов в каждом наборе. Все три оценки попадают в относительно узкий диапазон коэффициентов отбора: 0,002–0,02.
Характеристические коэффициенты отбора умеренно вредных мутаций: компьютерное моделирование
Основным недостатком описанного выше прямолинейного теоретического подхода является допущение, что эффективный размер человеческой популяции постоянен и равен 10 000 человек.Было показано, что это предположение подходит для обычных SNP, большинство из которых имеет древнее происхождение. Однако эффективный размер популяции, который на порядок больше, может быть более подходящим для описания динамики редких полиморфизмов. Большая эффективная численность популяции приводит к более эффективному очистительному отбору и, как видно из уравнения (1), к меньшим значениям R 1/ m ( s ). Независимым N E E , мы переоцениваем R 1/ M ( S ), и, на самом деле, R 1/ M ( S ) кривая зависимости от s смещена в область меньших значений коэффициентов отбора ().
Большая эффективная численность популяции приводит к более эффективному очистительному отбору и, как видно из уравнения (1), к меньшим значениям R 1/ m ( s ). Независимым N E E , мы переоцениваем R 1/ M ( S ), и, на самом деле, R 1/ M ( S ) кривая зависимости от s смещена в область меньших значений коэффициентов отбора ().
для анализа зависимости R 1/ M M ( S ) и R MAF > 0.25 ( S ) Под более реалистичным сценарием человеческой популяции. смоделированная эволюция по модели Райта-Фишера (при постоянной скорости мутаций и бесконечном количестве несцепленных сайтов). Наша модель истории человеческой популяции состояла из четырех основных эпох: ( a ) 100 000 поколений стабильной эффективной численности 15 000, ( b ) экспоненциальное сокращение за 100 поколений до размера 7000, ( c ) 500 поколения стабильного размера популяции 7000 и ( d ) экспоненциальный рост за 3000 поколений до эффективного размера популяции
.
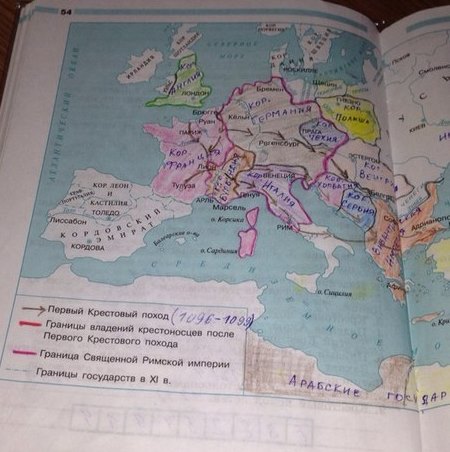 Такой сценарий, когда за продолжительным периодом стабильного фактического размера популяции следует узкое место, а затем быстрое увеличение, отражает ключевые черты истории человеческой популяции. Точные значения длительности каждой эпохи и соответствующие эффективные размеры популяции были выбраны в общем соответствии с последними литературными данными 41 – 43 , а затем были немного оптимизированы, чтобы лучше описать распределения частот аллелей для некодирующих SNP, обнаруженных с помощью NEIHS-ЕГП.Использование R 1/ M M ( S ) и R MAF > 0.25 ( S ) Зависимости, полученные с использованием прямого моделирования, привели к оценке, что ~ 0.001-0.003 Коэффициенты отбора соответствуют умеренно вредному классу миссенс-мутаций de novo.
Такой сценарий, когда за продолжительным периодом стабильного фактического размера популяции следует узкое место, а затем быстрое увеличение, отражает ключевые черты истории человеческой популяции. Точные значения длительности каждой эпохи и соответствующие эффективные размеры популяции были выбраны в общем соответствии с последними литературными данными 41 – 43 , а затем были немного оптимизированы, чтобы лучше описать распределения частот аллелей для некодирующих SNP, обнаруженных с помощью NEIHS-ЕГП.Использование R 1/ M M ( S ) и R MAF > 0.25 ( S ) Зависимости, полученные с использованием прямого моделирования, привели к оценке, что ~ 0.001-0.003 Коэффициенты отбора соответствуют умеренно вредному классу миссенс-мутаций de novo.
Кумулятивная равновесная частота умеренно вредных полиморфизмов в геноме человека
Мы определили, что умеренно вредные миссенс-мутации, не присутствующие среди обычных полиморфизмов, тем не менее многочисленны среди полиморфизмов с обнаруженной популяционной частотой <1% и связаны с коэффициентами отбора в диапазон 0. 001–0,003. Используя этот результат, мы оценили равновесную частоту таких умеренно вредных аллелей в человеческой популяции. В предположении сильного отбора и большого размера популяции ( N e × s ≫1) кумулятивная равновесная частота всех аллелей умеренно вредных генов в популяции составляет
001–0,003. Используя этот результат, мы оценили равновесную частоту таких умеренно вредных аллелей в человеческой популяции. В предположении сильного отбора и большого размера популяции ( N e × s ≫1) кумулятивная равновесная частота всех аллелей умеренно вредных генов в популяции составляет
, где u — частота мутаций. на ген на поколение, а s — коэффициент отбора, ассоциированный с гетерозиготами.Мы использовали 2×10 -8 как частоту мутаций на нуклеотид на поколение, рассчитанную Кондрашовым. 21 Важно отметить, что эта оценка кумулятивной частоты вредных аллелей не зависит от демографической истории населения.
Частота мутаций для умеренно вредных миссенс-мутаций на ген может быть оценена следующим образом: умеренно вредная миссенс-мутация составляет ~1%.
Принимая во внимание, что белок-кодирующие последовательности составляют ∼1,5% генома человека, мы подсчитали, что в геноме среднего человека присутствует ∼600 миссенс-мутаций с коэффициентом отбора в диапазоне 0,001–0,003. Следует отметить, что количество мутаций со значительно более слабым влиянием на приспособленность (со связанными с ними коэффициентами отбора <10 -4 ), вероятно, будет значительно выше.
Следует отметить, что количество мутаций со значительно более слабым влиянием на приспособленность (со связанными с ними коэффициентами отбора <10 -4 ), вероятно, будет значительно выше.
Обсуждение
Было отмечено, что при некоторых менделевских заболеваниях существуют менее тяжелые формы заболевания, связанные с более легкими мутациями, которые не полностью устраняют активность белка. 1 Было высказано предположение, что такие умеренно вредные миссенс-мутации могут служить генетической основой сложных заболеваний. Правдоподобие этой гипотезы зависит от того, насколько часты такие умеренно вредные замены среди мутаций de novo и насколько силен действующий на них очищающий отбор. О наличии вредных миссенс-полиморфизмов в геноме человека сообщалось во многих предыдущих исследованиях. 44 — — 55 — 55 В настоящей работе мы провели количественный анализ, рассчитав долю мягких вредных мутаций среди де Ново аминокислотные изменения и очень редкие и очень распространенные несинонимичные SNP. До недавнего времени отсутствие больших наборов данных повторного секвенирования, охватывающих десятки генов и сотни хромосом, препятствовало такому анализу, хотя в нескольких более ранних работах анализировалось распределение коэффициентов отбора для новых миссенс-мутаций у людей и мух с использованием зависимости скорости замещения от эффективной Размер населения, 56 — — 58 Сравнение полиморфизма и расходимости, 34 , 55 , 58 , 58 и частотный спектр в меньших образцах. 54 , 58 , 59
До недавнего времени отсутствие больших наборов данных повторного секвенирования, охватывающих десятки генов и сотни хромосом, препятствовало такому анализу, хотя в нескольких более ранних работах анализировалось распределение коэффициентов отбора для новых миссенс-мутаций у людей и мух с использованием зависимости скорости замещения от эффективной Размер населения, 56 — — 58 Сравнение полиморфизма и расходимости, 34 , 55 , 58 , 58 и частотный спектр в меньших образцах. 54 , 58 , 59
Мы рассчитали долю умеренно вредных миссенс-мутаций среди всех замен de novo как оставшуюся разницу после вычитания сильно вредных мутаций и эффективно нейтральных мутаций. Мы определили, что среди всех аминокислотных замен de novo сильно вредные мутации составляют около 20%. Полученное нами полногеномное значение аналогично доле аминокислотных замен, разрушающих функцию белка (25%), полученной Ямпольским и соавт. 34 Наша оценка также в целом согласуется с результатами Eyre-Walker et al., 54 , которые, подгоняя распределение коэффициентов отбора, смоделированное как гамма-функция, к данным о генетической изменчивости человека, оценили, что ∼15 % миссенс-замен являются сильно вредными, с коэффициентами отбора> 0,1. Ранее Фэй и соавт. 55 предположил, что 54% новых миссенс-мутаций у людей сильно вредны. Эта оценка относится к доле несинонимичных SNP, не наблюдаемых в выборке из 100 хромосом, поэтому эта оценка не может быть напрямую сопоставима с оценкой, представленной здесь.Наше значение доли эффективно нейтральных аминокислотных замен (27%) лишь немного выше предыдущей оценки 51 в 24% и соответствующей оценки K A K S 4 4 39 для сравнения человека и шимпанзе (23%).
Полученное нами полногеномное значение аналогично доле аминокислотных замен, разрушающих функцию белка (25%), полученной Ямпольским и соавт. 34 Наша оценка также в целом согласуется с результатами Eyre-Walker et al., 54 , которые, подгоняя распределение коэффициентов отбора, смоделированное как гамма-функция, к данным о генетической изменчивости человека, оценили, что ∼15 % миссенс-замен являются сильно вредными, с коэффициентами отбора> 0,1. Ранее Фэй и соавт. 55 предположил, что 54% новых миссенс-мутаций у людей сильно вредны. Эта оценка относится к доле несинонимичных SNP, не наблюдаемых в выборке из 100 хромосом, поэтому эта оценка не может быть напрямую сопоставима с оценкой, представленной здесь.Наше значение доли эффективно нейтральных аминокислотных замен (27%) лишь немного выше предыдущей оценки 51 в 24% и соответствующей оценки K A K S 4 4 39 для сравнения человека и шимпанзе (23%). Разница, скорее всего, связана с применением модели мутации, основанной на контексте. Меньшая предыдущая оценка 34 в 12%, возможно, была результатом использования слишком удаленной последовательности мыши в качестве внешней группы.
Разница, скорее всего, связана с применением модели мутации, основанной на контексте. Меньшая предыдущая оценка 34 в 12%, возможно, была результатом использования слишком удаленной последовательности мыши в качестве внешней группы.
На основании очень сильной разницы в соотношениях N a / N s для очень редких и очень распространенных миссенс-полиморфизмов мы подсчитали, что большинство редких миссенс-полиморфизмов обнаруживается в миссенс-полиморфизмах. человеческой популяции связаны с удивительно узким диапазоном коэффициентов отбора: 0,001–0,003. Из-за такого относительно мягкого очищающего отбора, действующего на них, они могут достигать высокой кумулятивной частоты в человеческой популяции, сохраняя при этом весьма гетерогенный спектр отдельных аллелей.
Увеличение отношения N a / N s было отмечено в другом месте 44 ; однако степень такой разницы не всегда была очевидна из-за небольшого числа секвенированных хромосом и грубого бинирования по частоте.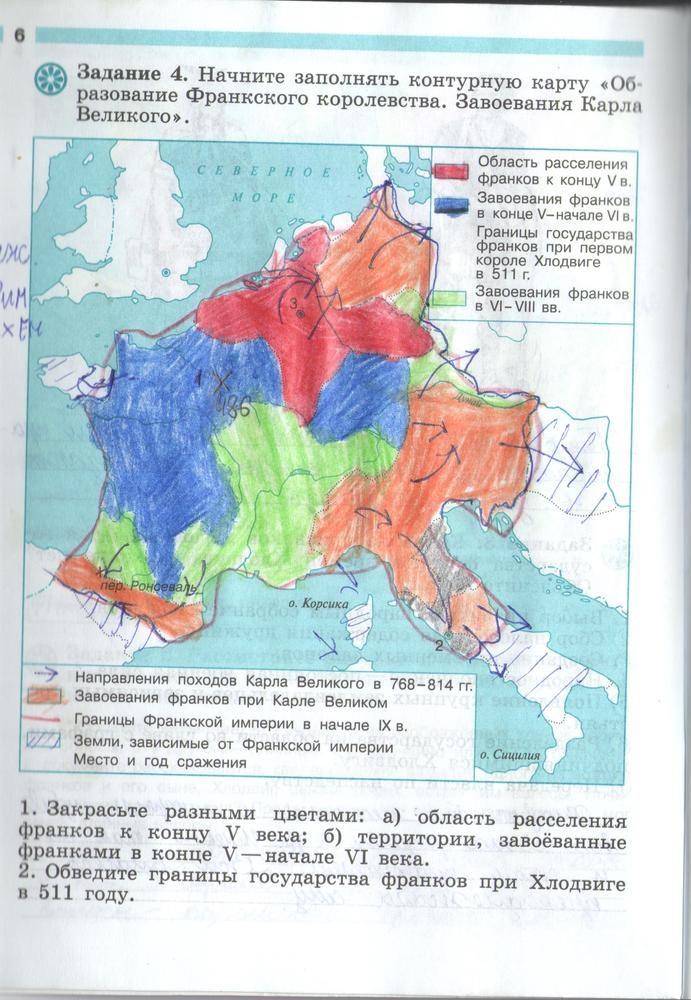 Сильное «пороговое» увеличение отношения N a / N s для SNP с частотой <6% было отмечено Wong et al. 60 Однако в этой работе 60 аналитически не рассматривалась возможность того, что наблюдаемый эффект является следствием формы распределения коэффициентов отбора для мутаций de novo.
Сильное «пороговое» увеличение отношения N a / N s для SNP с частотой <6% было отмечено Wong et al. 60 Однако в этой работе 60 аналитически не рассматривалась возможность того, что наблюдаемый эффект является следствием формы распределения коэффициентов отбора для мутаций de novo.
Мы подсчитали, что для среднего белка из 500 аминокислот кумулятивная равновесная частота аллелей, несущих вредные миссенс-SNP, примерно равна 1% в человеческой популяции. Если рассматривать необычно длинные белки или целые пути в качестве мишеней для мутаций, то частота их мутаций и, следовательно, равновесная частота умеренно вредных аллелей будет значительно выше 1%.На некоторые черты, такие как предрасположенность к сложным заболеваниям, могут влиять десятки, а может быть, и сотни генов. Мутационный объект для таких признаков будет исключительно большим. Большой размер мутационной мишени приводит к большой скорости мутаций на поколение и, как следствие, к большому уровню вредоносного полиморфизма, так как в равновесии он прямо пропорционален скорости мутаций. Даже для простого метаболического пути, который включает несколько мультисубъединичных ферментов, процент людей, имеющих хотя бы одну повреждающую мутацию в этом пути, может легко превысить 10%.Этот пример показывает, что общая частота редких умеренно вредных полиморфизмов достаточно высока, чтобы служить основанием для наследственной предрасположенности даже к большинству распространенных сложных заболеваний, таких как гипертония или ишемическая болезнь сердца. Это предполагает возможность того, что баланс мутаций и отбора может быть возможным эволюционным объяснением, по крайней мере, для некоторых распространенных заболеваний.
Даже для простого метаболического пути, который включает несколько мультисубъединичных ферментов, процент людей, имеющих хотя бы одну повреждающую мутацию в этом пути, может легко превысить 10%.Этот пример показывает, что общая частота редких умеренно вредных полиморфизмов достаточно высока, чтобы служить основанием для наследственной предрасположенности даже к большинству распространенных сложных заболеваний, таких как гипертония или ишемическая болезнь сердца. Это предполагает возможность того, что баланс мутаций и отбора может быть возможным эволюционным объяснением, по крайней мере, для некоторых распространенных заболеваний.
Исследования ассоциации генов-кандидатов
Эти результаты важны не только для нашего понимания роли редких несинонимичных SNP в предрасположенности к сложным заболеваниям, но также и для методов обнаружения генов, несущих в себе такие вредные генетические вариации.В настоящее время существует два основных подхода к идентификации генов, участвующих в комплексных заболеваниях: исследования сцепления 61 и ассоциативные исследования популяционных выборок. 61 , 62 В то время как первый подход является наиболее эффективным для идентификации менделевских редких генетических вариантов с высокой пенетрантностью, второй подход лучше подходит для идентификации распространенных вариантов с относительно низкой пенетрантностью. Однако ни один из этих подходов не способен обнаружить локусы восприимчивости, содержащие многочисленные, но редко встречающиеся умеренно вредные полиморфизмы.Однако наш анализ умеренно вредных мутаций показывает, что эта ситуация очень правдоподобна и может быть верной для многих генов и распространенных заболеваний.
61 , 62 В то время как первый подход является наиболее эффективным для идентификации менделевских редких генетических вариантов с высокой пенетрантностью, второй подход лучше подходит для идентификации распространенных вариантов с относительно низкой пенетрантностью. Однако ни один из этих подходов не способен обнаружить локусы восприимчивости, содержащие многочисленные, но редко встречающиеся умеренно вредные полиморфизмы.Однако наш анализ умеренно вредных мутаций показывает, что эта ситуация очень правдоподобна и может быть верной для многих генов и распространенных заболеваний.
В свете наших результатов недавно предложенный альтернативный подход выглядит более многообещающим. Этот подход направлен на обнаружение обогащения редкими, потенциально вредными миссенс SNP в группе пациентов по сравнению с контрольной группой. 63 , 64 Рассматривая совокупную частоту вредных мутаций, а не их индивидуальную частоту, этот метод подходит для исследования распространенных заболеваний, которые имеют очень гетерогенный спектр предрасполагающих аллелей. Такой миссенс-аккумуляционный подход к исследованиям ассоциации генов-кандидатов был недавно успешно использован в исследованиях гена MC4R , 65 тирозинфосфатома при колоректальном раке, 66 и генов, участвующих в нарушениях липидного обмена. 67
Такой миссенс-аккумуляционный подход к исследованиям ассоциации генов-кандидатов был недавно успешно использован в исследованиях гена MC4R , 65 тирозинфосфатома при колоректальном раке, 66 и генов, участвующих в нарушениях липидного обмена. 67
Однако были выявлены два основных потенциальных недостатка этого метода. 63 Во-первых, это высокая стоимость полного геномного секвенирования генов-кандидатов у большого числа людей.Во-вторых, это аналитическая задача выбора потенциально вредных SNP из большого количества нейтральных генетических вариаций. 63 Первое возражение против этого метода почти наверняка будет устранено в ближайшем будущем благодаря продолжающемуся резкому сокращению затрат на секвенирование 68 и разработке новых стратегий секвенирования. 69 Второе возражение является более фундаментальным, поскольку неспособность различить вредные и нейтральные аминокислотные замены привела бы к очень низкому соотношению сигнал/шум. Чтобы изучить обогащение вредными мутациями, нужно уметь обнаруживать их среди нейтральной генетической изменчивости. Ранее считалось, что количество вредных аллелей в фенотипах заболеваний невелико по сравнению с большим количеством нейтральных миссенс-SNP, присутствующих как в группах больных, так и в контрольных группах. Наш анализ, однако, показывает, что большинство миссенс-замен с обнаруженной частотой <1% на самом деле вредны. Таким образом, если учитывать только замены с выявленной частотой <1%, то обогащение числа мутаций в группе заболеваний должно быть сильно выражено.
Чтобы изучить обогащение вредными мутациями, нужно уметь обнаруживать их среди нейтральной генетической изменчивости. Ранее считалось, что количество вредных аллелей в фенотипах заболеваний невелико по сравнению с большим количеством нейтральных миссенс-SNP, присутствующих как в группах больных, так и в контрольных группах. Наш анализ, однако, показывает, что большинство миссенс-замен с обнаруженной частотой <1% на самом деле вредны. Таким образом, если учитывать только замены с выявленной частотой <1%, то обогащение числа мутаций в группе заболеваний должно быть сильно выражено.
Выводы
Большая доля умеренно вредных мутаций среди миссенс-мутаций предполагает, что баланс мутации-отбора является правдоподобным объяснением существования распространенного заболевания со сложным наследованием, по крайней мере, в некоторых случаях. Кроме того, вполне возможно, что некоторые распространенные заболевания могут быть вызваны множеством редких аллельных вариантов. Наблюдение, что большинство редких несинонимичных вариантов человека являются вредными и, таким образом, имеют значение для функции и фенотипа, предлагает стратегию исследований ассоциации генов-кандидатов. Ожидается, что больные популяции будут иметь более высокий уровень редких аминокислотных вариантов в генах, участвующих в заболевании, чем здоровые контрольные популяции. Эту разницу можно легко обнаружить в ходе глубокого повторного секвенирования. Очевидно, что эта стратегия была бы крайне неэффективной, если бы большинство вариантов кодирования на низкой частоте были нейтральными. Несколько недавних сообщений продемонстрировали избыток редких миссенс-вариантов у людей с фенотипами, связанными с риском заболевания. Наш анализ объясняет успех этих исследований.
Ожидается, что больные популяции будут иметь более высокий уровень редких аминокислотных вариантов в генах, участвующих в заболевании, чем здоровые контрольные популяции. Эту разницу можно легко обнаружить в ходе глубокого повторного секвенирования. Очевидно, что эта стратегия была бы крайне неэффективной, если бы большинство вариантов кодирования на низкой частоте были нейтральными. Несколько недавних сообщений продемонстрировали избыток редких миссенс-вариантов у людей с фенотипами, связанными с риском заболевания. Наш анализ объясняет успех этих исследований.
Крюков канал в Санкт-Петербурге
Этот исторический канал менее известен, чем петербургская тройка узких центральных водных путей — канал Грибоедова и реки Мойка и Фонтанка. Однако он проходит перпендикулярно и соединяет все три из них. Его отсутствие признания является позором. Во-первых, потому, что его гранитные набережные и старые мосты довольно красивы, а во-вторых, потому, что на его берегах стоят многие из самых впечатляющих достопримечательностей Санкт-Петербурга, такие как Новая Голландия, Мариинский театр и колокольня собора Святого Николая. Никольский морской собор. История канала также во многом уникальна.
Никольский морской собор. История канала также во многом уникальна.
Панорама Крюкова канала
Крюков канал первоначально был прорыт от Невы до реки Мойки в 1717-1720 гг. Руководил работами некий Семен Крюков, в честь которого и был назван канал. Затем, в 1782-1787 годах, его продлили до реки Фонтанки. В начале 19 века канал был оборудован гранитными насыпями, а в 1847 году участок от Невы до Благовещенской площади (Благовещенская площадь, ныне площадь Труда) был проложен по трубе, что позволило выйти к Неве. только через Адмиралтейский и Ново-Адмиралтейский каналы.
Одной из главных достопримечательностей Крюкова канала является рукотворный остров Новая Голландия (Новая Голландия). Он появился в момент начала рытья канала, а в 1765-1780 годах остров был застроен краснокирпичными складами под руководством Саввы Чевакинского. Его вход в водный путь украшала монументальная неоклассическая арка из красного кирпича, спроектированная Жаном-Батистом Валленом де ла Мотом в 1788 году. Параллельно складам на противоположном берегу канала стоят массивные Крюковские морские казармы.
Параллельно складам на противоположном берегу канала стоят массивные Крюковские морские казармы.
Вид на Мариинский театр с набережной Крюкова канала
Вдоль набережной к югу от Новой Голландии с конца 18 века стоял Литовский замок. Это своеобразное здание в виде башен неправильного пятиугольника было спроектировано Иваном Старовым в 1787 году для размещения тюрьмы. Он оставался пустым более десяти лет после постройки, а затем был передан в качестве казармы Литовскому мушкетерскому полку, отсюда и его название. В 1820-х годах он был преобразован в тюрьму и использовался для заключения жестоких преступников и бродяг, а также некоторых политических заключенных.Радикальный писатель Владимир Кораленко был заключен здесь в 1870-х годах за распространение революционных брошюр, а известный (и известный алкоголик) писатель Александр Куприн был задержан в октябре 1903 года за пьяную драку в ресторане. Во время Февральской революции 1917 года у тюрьмы собралась толпа, и она была сожжена дотла. Его фундамент позже был использован при строительстве импозантного сталинского жилого дома Союза рабочих причалов.
Его фундамент позже был использован при строительстве импозантного сталинского жилого дома Союза рабочих причалов.
Рядом с Литовским замком через улицу Декабристов и построенный одновременно с замком Литовский рынок был одним из нескольких торговых рядов, спроектированных Джакомо Кваренги, похожих по дизайну на его Малый гостиный двор на Думской улице и тем более на Никольский. Рынок дальше на восток вдоль канала.К 1920-м годам рынок пришел в упадок и после пожара на его месте разместился конструктивистский Дворец культуры первой пятилетки. Дворец был популярной культурной площадкой, где выступали многие известные советские артисты и музыкальные коллективы. Как ни странно, в 2005 году это массивное здание было снесено для строительства второй сцены Мариинского театра. Мариинский-2, открытый в конце сезона 2013 года, является самым современным зданием Крюкова канала, обращенным к задней части главного Мариинского театра через канал.
Лед на Крюковом канале
В 1855-1861 годах через канал от Литовского замка был построен доходный дом барона Фитингова. Строительством руководил шведско-российский архитектор Карл Андерсон. Сегодня это музейно-выставочный центр «Петербургский художник».
Строительством руководил шведско-российский архитектор Карл Андерсон. Сегодня это музейно-выставочный центр «Петербургский художник».
Одним из самых ярких зданий вдоль канала является жилой дом «Веге» (№ 14) на углу проспекта Римского-Корсаково. Построенный для владельца шоколадной фабрики в 1912-1914 годах Сергеем Овсянниковым в стиле неоклассицизма, темно-серый гранитный фасад здания богато украшен скульптурой и высшими рельефами, а центральная арка поддерживается могучими атлантами.При входе в здание посетителей встречает статуя Пана, отдыхающего со своей трубкой на скамейке в вестибюле.
Санкт-Петербург — Северная Венеция
На противоположном берегу канала, в доме № 15, построено величественное здание в стиле неоклассицизма более ранней эпохи для размещения Гимназии Императорского благотворительного общества (№ 15). Первоначальная гимназия была построена двумя лучшими архитекторами того времени, Константином Тоном и Василием Стасовым.Гимназия стоит напротив, пожалуй, самого красивого здания вдоль канала, Морского собора Святого Николая. Этот красивый собор в стиле барокко, построенный между 1753 и 1762 годами Саввой Чевакинским, стоит в стороне от канала, с высокой колокольней у самой воды.
Этот красивый собор в стиле барокко, построенный между 1753 и 1762 годами Саввой Чевакинским, стоит в стороне от канала, с высокой колокольней у самой воды.
Шесть мостов пересекают канал. Все мосты, кроме Кашина, представляют собой трехпролетные каменные мосты, облицованные гранитом. Мосты неоднократно перестраивались и реконструировались, но сохранили свой первоначальный вид.Из сохранившихся декоративных элементов на мостах стоит отметить оригинальные кованые решетки на Старо-Никольский мост и Декабристский мост и металлические фонарные столбы Торгового и Матвеева мостов. Как известно всем, кто путешествует по Крюкову каналу на лодке, Старо-Никольский мост также является местом, с которого можно увидеть больше мостов, чем где-либо еще в Санкт-Петербурге — всего семь вдоль Крюкова канала и канала Грибоедова. .
| Характеристики реки: | Длина: 1.015 км |
|---|---|
| Основные мосты: |
Смежный мост, Старо-Никольский мост, Кашин мост, Торговый мост, Мост Декабристов, Матвеев мост |
ОТЕЛИ: Выбор редакции
ОБЩЕЖИТИЯ: Выбор редакции
РЕСТОРАНЫ: Выбор редакции
Демографическая реконструкция древней ДНК подтверждает быстрое исчезновение бескрылой гагарки
Основные версии:
1) Исследовательские предположения:
Рецензенты отметили, что некоторые аспекты исследования неизбежно оставались весьма спекулятивными, и сочли, что это следует лучше признать во всем.
Поэтому, пожалуйста, четко укажите предположения и степень достоверности для всех результатов и выводов; вся ключевая информация должна содержаться в основном тексте, а не только в онлайн-приложении.
Спасибо за отзыв. Там, где это уместно, мы попытались улучшить ясность наших предположений, степень достоверности различных результатов и выводов. В частности, мы более подробно обсудили наши выводы о быстром сокращении населения и о том, что показывают и не показывают наши данные (первый абзац Обсуждения).Мы также более четко выявили спекуляции при обсуждении данных GPS. Кроме того, мы добавили моделирование вихрей с «реалистичными» показателями смертности и воспроизводства, полученными от гагарки, чтобы поместить наши консервативные оценки устойчивого вылова в контекст и подчеркнуть неопределенность наших оценок. Наконец, по просьбе рецензентов мы переместили рисунок TempNet в основной текст.
2) Капсулы GPS Drift:
По общему мнению, это был один из самых слабых аспектов исследования.
В частности, неясно, можно ли достоверно нанести на карту токи всего с двумя капсулами и можно ли использовать эти данные для надежных выводов о течениях тысячелетней давности. Кроме того, предполагается ли, что течения стабильны в течение всего года? Взрослые бескрылые гагарки, по-видимому, перемещались только в определенное время года, и, поскольку они почти наверняка были филопатриками, однонаправленные стрелки на карте поднимают вопрос о том, как они возвращались в свои колонии. Пожалуйста, смягчите выводы, сделанные на основе набора данных дрейфующей капсулы, выделите моделирование океанских течений как задачу для будущей работы и предоставьте лучшее обоснование для этого направления исследования (бескрылая гагарка была нелетающей птицей и, следовательно, могла полагаться или, по крайней мере, подвержены влиянию океанских течений).
Мы внесли поправки в несколько разделов рукописи, в которых обсуждаются данные капсулы GPS. Внесены следующие изменения:
Результаты: Данные об океанских течениях. Чтобы оценить потенциальные причины наблюдаемого отсутствия структуры популяции, мы получили данные с дрейфующих капсул с GPS, которые были развернуты в Северной Атлантике в рамках проекта «Послание в бутылке» Verkís Consulting. Инженеры. Поскольку бескрылая гагарка не летала, океанские течения могли повлиять на характер ее миграции.Маршрут, пройденный капсулами, соединяет некоторые из основных гнездовых колоний в Сент-Килде (Шотландия), Гейрфугласкер/Элдей-Айленд (Исландия) и Фанк-Айленд (Канада) (рис. 4). Экстраполяция современных данных об океанских течениях в прошлое и интерпретация этих данных в контексте перемещений больших гагарок носят чисто спекулятивный характер. Однако, если современные океанские течения сопоставимы с течениями прошлых тысячелетий, то эти данные, по крайней мере, дают возможное объяснение того, как бескрылые гагарки перемещались через свой прежний ареал и между гнездовыми колониями (рис. 4).Полное описание маршрутов движения капсул приведено в Приложении 1.
Чтобы оценить потенциальные причины наблюдаемого отсутствия структуры популяции, мы получили данные с дрейфующих капсул с GPS, которые были развернуты в Северной Атлантике в рамках проекта «Послание в бутылке» Verkís Consulting. Инженеры. Поскольку бескрылая гагарка не летала, океанские течения могли повлиять на характер ее миграции.Маршрут, пройденный капсулами, соединяет некоторые из основных гнездовых колоний в Сент-Килде (Шотландия), Гейрфугласкер/Элдей-Айленд (Исландия) и Фанк-Айленд (Канада) (рис. 4). Экстраполяция современных данных об океанских течениях в прошлое и интерпретация этих данных в контексте перемещений больших гагарок носят чисто спекулятивный характер. Однако, если современные океанские течения сопоставимы с течениями прошлых тысячелетий, то эти данные, по крайней мере, дают возможное объяснение того, как бескрылые гагарки перемещались через свой прежний ареал и между гнездовыми колониями (рис. 4).Полное описание маршрутов движения капсул приведено в Приложении 1.
Обсуждение: Мы можем только догадываться, какие факторы могли привести к такому отсутствию структуры популяции, но данные, собранные с дрейфующих капсул с GPS, согласуются с гипотезами, выдвинутыми рядом авторов. Было высказано предположение, что миграции происходили как в северном, так и в южном направлениях между местами размножения и зимовки, чему способствовали океанические течения, такие как Восточно-Гренландское течение (Brown, 1985; Meldgaard, 1988; Montevecchi and Kirk, 1996).Однако, поскольку эти предварительные данные были доступны только от двух дрейфующих капсул с GPS, а океанские течения могли значительно измениться за последние несколько столетий, выводы, которые мы можем сделать из таких данных, несколько ограничены. Кроме того, возможно, что эти токи могут меняться в течение года. Таким образом, к этим данным следует относиться с осторожностью в ожидании гораздо более подробных исследований океанских течений в Северной Атлантике в течение года. Тем не менее высокая изменчивость бескрылой гагарки дополнительно подтверждается ее способностью отслеживать свою среду обитания в ответ на изменение климата, о чем свидетельствуют археологические данные (Bengtson, 1984; Campmas et al., 2010; Мелдгаард, 1988; Сержантсон, 2001).
Было высказано предположение, что миграции происходили как в северном, так и в южном направлениях между местами размножения и зимовки, чему способствовали океанические течения, такие как Восточно-Гренландское течение (Brown, 1985; Meldgaard, 1988; Montevecchi and Kirk, 1996).Однако, поскольку эти предварительные данные были доступны только от двух дрейфующих капсул с GPS, а океанские течения могли значительно измениться за последние несколько столетий, выводы, которые мы можем сделать из таких данных, несколько ограничены. Кроме того, возможно, что эти токи могут меняться в течение года. Таким образом, к этим данным следует относиться с осторожностью в ожидании гораздо более подробных исследований океанских течений в Северной Атлантике в течение года. Тем не менее высокая изменчивость бескрылой гагарки дополнительно подтверждается ее способностью отслеживать свою среду обитания в ответ на изменение климата, о чем свидетельствуют археологические данные (Bengtson, 1984; Campmas et al., 2010; Мелдгаард, 1988; Сержантсон, 2001).
Мы надеемся, что теперь это проясняет, что, хотя данные были включены в Обсуждение и использованы для поддержки некоторых наших выводов, они взяты из небольшого предварительного набора данных и поэтому должны рассматриваться именно так.
3) Геномный анализ:
Рецензенты подняли несколько вопросов, касающихся геномного анализа, которые требуют решения.
а) Есть намек на то, что испанская выборка могла отличаться от северной и могла представлять рефугиальное население.Были ли какие-либо частичные данные мтДНК из других 8 образцов из Испании, которые можно было бы использовать для дальнейшего изучения?
Спасибо за это замечание. Мы изучили это как можно лучше с ограниченными данными (см. Ниже). Наши результаты показывают, что в Испании нет никаких доказательств существования рефугиального населения. Образцы из Испании так же распределены по дереву, как и образцы из других регионов. Ниже мы суммировали дополнительные анализы, которые мы провели. Однако, поскольку они не показывают какой-либо закономерности, не наблюдаемой в данных, которые мы используем для этой публикации, мы бы предпочли не включать этот раздел в рукопись.Мы считаем, что это повлияет на поток и ясность рукописи без добавления какой-либо важной информации. Мы, конечно, готовы включить этот раздел, например, в приложение, если редактор сочтет это целесообразным.
Однако, поскольку они не показывают какой-либо закономерности, не наблюдаемой в данных, которые мы используем для этой публикации, мы бы предпочли не включать этот раздел в рукопись.Мы считаем, что это повлияет на поток и ясность рукописи без добавления какой-либо важной информации. Мы, конечно, готовы включить этот раздел, например, в приложение, если редактор сочтет это целесообразным.
Дополнительные анализы Испанские образцы.
В филогенетическом дереве бескрылых бескрылых, которое дало достаточно данных о последовательностях в соответствии с нашими критериями фильтрации (Приложение 4), образец MK40_Spain оказался дифференцированным от остальных образцов, происходящих из северных районов их распространения.В связи с этим возникает вопрос, может ли выборка представлять население-беженцы в Испании. Чтобы проверить это, мы повторно изучили филогенетические отношения между образцами с добавлением других испанских образцов, которые мы секвенировали, но которые не соответствовали критериям фильтрации для включения в наш окончательный набор данных. Эти образцы включали MK37, MK42, MK44 и MK45. Эти образцы характеризовались плохим покрытием (средний охват варьировался от 0,18 до 2,07) и более чем 33% оснований, отсутствующих в консенсусной последовательности (длина консенсусной последовательности варьировалась от 36 п.н. до 5468 п.н.).Последовательности, созданные, как описано, с использованием пайплайна Paleomix (Приложение 4) для образцов MK37, MK42, MK44 и MK45, были вручную сопоставлены с эталонным геномом с использованием Bioedit v7.2.5 (Hall, 1999) и Tablet v1.16.09.06 (Milne et al. , 2013), чтобы просмотреть масштабированную двоичную карту выравнивания (BAM). Поскольку MK37 и MK45 были очень низкого качества, мы не смогли использовать их в этом дополнительном анализе. Однако мы смогли произвести выравнивание 859 п.н., которое включало дополнительные испанские образцы MK42 и MK44. Анализ соседнего соединения выравнивания на основе p-расстояний (Saitou and Nei, 1987) с использованием MEGAX (Kumar et al., 2018) дал очень плохо решенную филогению (ответ автора, изображение 1).
Эти образцы включали MK37, MK42, MK44 и MK45. Эти образцы характеризовались плохим покрытием (средний охват варьировался от 0,18 до 2,07) и более чем 33% оснований, отсутствующих в консенсусной последовательности (длина консенсусной последовательности варьировалась от 36 п.н. до 5468 п.н.).Последовательности, созданные, как описано, с использованием пайплайна Paleomix (Приложение 4) для образцов MK37, MK42, MK44 и MK45, были вручную сопоставлены с эталонным геномом с использованием Bioedit v7.2.5 (Hall, 1999) и Tablet v1.16.09.06 (Milne et al. , 2013), чтобы просмотреть масштабированную двоичную карту выравнивания (BAM). Поскольку MK37 и MK45 были очень низкого качества, мы не смогли использовать их в этом дополнительном анализе. Однако мы смогли произвести выравнивание 859 п.н., которое включало дополнительные испанские образцы MK42 и MK44. Анализ соседнего соединения выравнивания на основе p-расстояний (Saitou and Nei, 1987) с использованием MEGAX (Kumar et al., 2018) дал очень плохо решенную филогению (ответ автора, изображение 1). Важно отметить, что новые испанские образцы не группируются с выбросом MK40, тем самым не подтверждая гипотезу об испанском рефугиальном населении.
Важно отметить, что новые испанские образцы не группируются с выбросом MK40, тем самым не подтверждая гипотезу об испанском рефугиальном населении.
Эволюционная история была выведена с использованием метода Neighbor-Joining [Saitou and Nei, 1987].
Оптимальное дерево с суммой длин ветвей = 0.01164144 показан. Рядом с ветвями показан процент повторяющихся деревьев, в которых ассоциированные таксоны сгруппированы вместе в тесте начальной загрузки (1000 повторов) [Felsenstein, 1985]. Дерево нарисовано в масштабе с длинами ветвей в тех же единицах, что и эволюционные расстояния, используемые для вывода филогенетического дерева. Эволюционные расстояния были рассчитаны с использованием метода p-расстояний [Nei and Kumar, 2000] и выражены в единицах количества базовых различий на сайт. В этом анализе участвовали 43 нуклеотидные последовательности. Все позиции, содержащие пробелы и отсутствующие данные, были устранены (опция полного удаления). Всего в окончательном наборе данных было 859 позиций. Эволюционный анализ проводился в MEGA X [Kumar et al., 2018].
Все позиции, содержащие пробелы и отсутствующие данные, были устранены (опция полного удаления). Всего в окончательном наборе данных было 859 позиций. Эволюционный анализ проводился в MEGA X [Kumar et al., 2018].
б) К сожалению, образцы с острова Фанк не имели даты и были исключены из анализов, хотя их добавление с большим априором мало повлияло на результаты; пожалуйста, добавьте соответствующее дерево в онлайн-приложение (например, Приложение — рис. 4).
Мы согласны с тем, что, к сожалению, образцы с острова Фанк не имеют информации о дате, однако, к сожалению, такова природа образца из этого места, которое, по сути, представляет собой братское кладбище бескрылых гагарок с сотнями костей периода интенсивная охота. Конечно, в будущем было бы интересно провести углеродную датировку некоторых образцов с острова Фанк, чтобы добавить эту информацию к набору данных. Однако на данный момент мы добавили все недатированные образцы в дерево и добавили его в дополнение по запросу (Новое приложение — рис. 5).
5).
Нет ли дополнительных исторических музейных образцов или образцов перьев, которые могли бы дать больше и более качественные данные о последовательности, чем старые субфоссильные кости?
Это исследование было частью проекта PhD, который был завершен в 2017 году, поэтому, к сожалению, финансирование и персонал для облегчения дальнейшего отбора проб больше не доступны. В ходе проекта была проведена обширная выборка, отбирая образцы, которые представляли особей из как можно большей части бывшего ареала бескрылой гагарки и в течение максимально возможного периода времени.Образцы были собраны из ряда музеев по всему миру. Поскольку мы хотели включить в набор данных образцы, содержащие как можно больше информации, мы решили не включать многие смонтированные музейные образцы, поскольку подавляющее большинство из них не имеют даты или информации о местоположении образцов.
в) Удивляет полное отсутствие филогеографической структуры.
Такая неструктурированность обычно объясняется либо высокой дисперсией, либо относительно недавним узким местом с последующим расширением ареала.Поскольку бескрылая гагарка была нелетающей птицей, было бы очень интересно, если бы вы могли добавить сравнение с филогеографической структурой (или ее отсутствием), обнаруженной у пингвинов (например, Обсуждение, третий абзац).
Спасибо за предложение сравнить наш результат с пингвинами. Это помогает поместить наши выводы, сделанные в отношении бескрылой гагарки, в контекст и показывает, что, хотя отсутствие структуры популяции бескрылой гагарки вызывает удивление, на самом деле это не кажется чем-то необычным для нелетающих морских птиц.Теперь мы добавили абзац, который гласит: «Хотя все ближайшие родственники бескрылой гагарки способны летать, что способствовало бы связности популяции, аналогичным образом сообщалось об отсутствии структуры популяции у некоторых видов пингвинов». Например, о структуре популяции императорского пингвина ( Aptenodytes forsteri ) (Cristofari et al.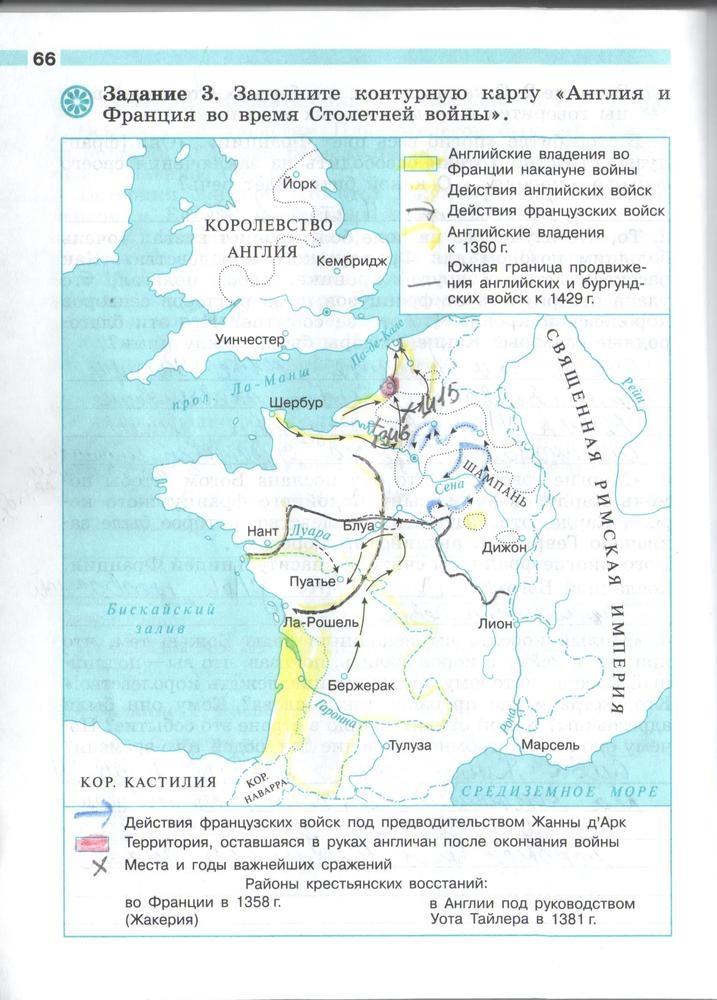 , 2016), антарктического пингвина ( Pygoscelis antarcticus) (Mura-Jornet et al., 2018) и Пингвин Адели ( стр.adeliae ) Gorman et al., 2017; Родер и др., 2001).
, 2016), антарктического пингвина ( Pygoscelis antarcticus) (Mura-Jornet et al., 2018) и Пингвин Адели ( стр.adeliae ) Gorman et al., 2017; Родер и др., 2001).
d) В связи с предыдущим комментарием, рукопись может быть улучшена кратким сравнением, возможно, в Обсуждении, с тем, что было обнаружено в предыдущих исследованиях при анализе данных мтДНК, отобранных во времени, в периоды демографического спада (например, либо у вымерших видов, либо у видов, которые находятся на грани исчезновения). сегодня находится под угрозой исчезновения). Рукопись, по-видимому, не цитирует какие-либо документы, в которых анализировались генетические изменения в рассматриваемых здесь временных масштабах (т.э., несколько столетий). Возможно, вызванное деятельностью человека сокращение численности бескрылой гагарки было даже более быстрым, чем сокращение численности многих видов, наблюдавшееся в прошлом столетии? Достаточно ли у вас образцов (то есть мощности), чтобы обнаружить снижение генетического разнообразия за последние 100 лет до вымирания бескрылой гагарки?
Это действительно было очевидным упущением в нашем Обсуждении. Теперь мы добавили раздел к первому абзацу Обсуждения, в котором обсуждался этот момент следующим образом:
Теперь мы добавили раздел к первому абзацу Обсуждения, в котором обсуждался этот момент следующим образом:
Если бы бескрылая гагарка находилась под угрозой исчезновения до начала интенсивной охоты человека, например, в результате длительной неоптимальной среды обитания или изменения окружающей среды, мы ожидали бы увидеть генетические доказательства такого стресса, как, например, наблюдаемое в исследованиях пещерных медведей (Stiller et al., 2010) и бизонов (Шапиро и др., 2004). С другой стороны, если бы популяция быстро сокращалась, например, в результате обширной охоты, генетические данные имели бы лишь очень ограниченную возможность обнаружить такое сокращение у долгоживущих видов. Исследования митохондриальной ДНК новозеландских моа не обнаружили признаков сокращения популяции до вымирания (Allentoft et al., 2014; Rawlence et al., 2012), и исследование эндемичных гавайских буревестников пришло к аналогичному выводу (Welch et al. , 2012). На самом деле, даже недавнее полногеномное исследование двух вымерших новозеландских певчих птиц (гуйя и кыкако с Южного острова), которые исчезли после заселения людьми в течение 700 лет, не обнаружило генетических доказательств сокращения популяции до исчезновения вида (Dussex et al. др., 2019). Таким образом, наши результаты согласуются с быстрым сокращением численности бескрылых гагарок. Однако важно иметь в виду, что наши результаты просто указывают на то, что гибель бескрылой гагарки вышла за пределы обнаружения генетических данных. Они не обязательно подтверждают, началась ли быстрая гибель, которая должна была произойти до вымирания, до или после начала экстенсивной охоты человека, а также не дают указаний на то, произошло ли более чем одно сокращение популяции. Локализованное, необъяснимое сокращение численности бескрылой гагарки на восточной стороне Северной Атлантики за последние 2000 лет, например, вытекает из сокращения находок костей в Англии, Шотландии и Скандинавии (Bengtson, 1984; Best and Mulville, 2014; Grieve, 1885; Hufthammer, 1982; Serjeantson, 2001), по-видимому, не были достаточно серьезными, чтобы оставить генетическую подпись.
др., 2019). Таким образом, наши результаты согласуются с быстрым сокращением численности бескрылых гагарок. Однако важно иметь в виду, что наши результаты просто указывают на то, что гибель бескрылой гагарки вышла за пределы обнаружения генетических данных. Они не обязательно подтверждают, началась ли быстрая гибель, которая должна была произойти до вымирания, до или после начала экстенсивной охоты человека, а также не дают указаний на то, произошло ли более чем одно сокращение популяции. Локализованное, необъяснимое сокращение численности бескрылой гагарки на восточной стороне Северной Атлантики за последние 2000 лет, например, вытекает из сокращения находок костей в Англии, Шотландии и Скандинавии (Bengtson, 1984; Best and Mulville, 2014; Grieve, 1885; Hufthammer, 1982; Serjeantson, 2001), по-видимому, не были достаточно серьезными, чтобы оставить генетическую подпись.
e) Дополнительный рисунок S4: добавьте версию рисунка TempNet с более высоким разрешением в качестве основного рисунка, а не в качестве дополнительного элемента.
Причина этого отчасти в том, что в настоящее время в основном тексте нет легкодоступной информации о том, как выборки распределены во времени, что важно для контекста. Более того, вывод о том, что разнообразие в последнем интервале времени (<250 лет) все еще было высоким, очень интересен и должен быть обсужден более подробно.
Мы добавили изображение рисунка TempNet с более высоким разрешением и переместили его в основной текст (рис. 2). Мы надеемся, что разрешение изображения теперь соответствует вашим требованиям.
4) Анализ жизнеспособности популяции:
Рецензенты в целом сочли анализ жизнеспособности популяции (PVA) хорошо реализованным, но выделили несколько моментов, требующих решения.
а) Вы выбрали довольно оптимистичный сценарий истории жизни бескрылой гагарки, полагая, что, если модель предсказывает вымирание даже в этих условиях, можно с уверенностью заключить, что риск исчезновения в реальном мире должен быть высоким.
Хотя это разумный подход, настройки PVA были, возможно, слишком оптимистичными, с нулевыми экологическими вариациями в размножении и очень низкими вариациями в уровне смертности (устанавливается на уровне 1% для всех возрастных классов).
Чтобы предоставить некоторые сравнительные данные с более «реалистичными» настройками, мы теперь добавили моделирование с показателями смертности, полученными от гагарки (включая более высокие стандартные вариации), увеличенным репродуктивным возрастом (с 4 до 5 лет) и уменьшенным максимальным репродуктивным возрастом. (до 20 лет с 25) См. Дополнительный файл 2B).Эти симуляции обеспечивают устойчивый уровень вылова около 40 000 птиц в год без сбора яиц. Вероятно, это недооценка, поскольку у гагарки действительно более высокая репродуктивная способность, чем у гагарки, и поэтому она может терпеть более высокую смертность. Это моделирование и его результаты теперь представлены и обсуждаются в соответствующих разделах рукописи.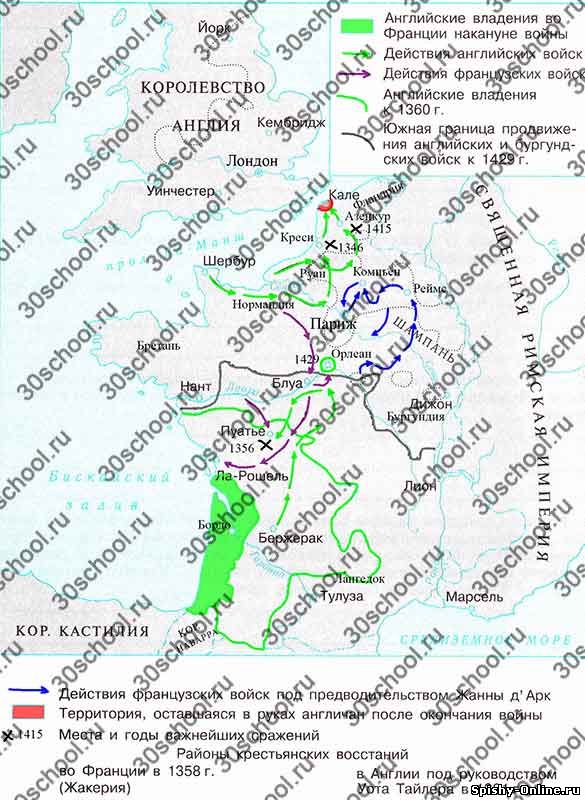
Более того, кажется, что Vortex работал с нормой вылова, установленной как постоянный процент от размера популяции (пожалуйста, уточните в редакции, так ли это было на самом деле, так как это было трудно сказать ни по рукописи, ни по документации Vortex), даже хотя вполне возможно, что люди могли продолжать ловить постоянное количество (по крайней мере, какое-то время), поскольку было бы относительно легко найти птиц в их гнездовых колониях.При запуске с такими базовыми настройками PVA приближается к детерминированной модели, и простой расчет λ мог бы дать такое же понимание: используя параметры истории жизни из дополнительных рис. 2A и B, λ можно оценить как ~ 1,12, а один пришли бы к выводу, что уровень вылова в 12% будет явно неустойчивым в детерминированном виде.
Извините, это не было хорошо описано в нашей рукописи. На самом деле мы использовали постоянные числа, а не постоянный процент.Теперь мы разъяснили это, где это уместно, на протяжении всей рукописи.
Рассмотрите возможность изучения более реалистичного набора настроек и предоставьте дополнительный контекст для обоснования вашего подхода к PVA.
b) В целом, параметры истории жизни тщательно подобраны, но немного странно, что ежегодная смертность молоди (13%) выше, чем смертность птенцов/птенцов (9%). Можно ли это оправдать или скорректировать?
Причина несколько странного распределения коэффициентов смертности заключалась в том, что мы хотели получить строгий способ определения коэффициентов в отсутствие биологической информации.Следовательно, мы выбрали критерий «самые низкие показатели из всех алцидов, обнаруженных в литературе». И самые низкие ставки, которые мы могли найти для периода 0-1 года, ниже, чем для более поздних периодов времени. Мы могли бы увеличить значения 0-1, но это было бы произвольно, поэтому мы отказались от этого. Тем не менее, теперь мы также включили анализы с полным набором уровней смертности гагарок и более высокими стандартными вариациями, чтобы обеспечить сравнение того, как наши симуляции будут выглядеть с менее консервативными, но все же зависящими от вида настройками. Мы также добавили дополнительное объяснение наших рассуждений о консервативных значениях в рукопись, которая гласит (Приложение 7):
Мы также добавили дополнительное объяснение наших рассуждений о консервативных значениях в рукопись, которая гласит (Приложение 7):
«Строгое применение правила, согласно которому мы используем самый низкий уровень смертности среди всех алцидных видов, найденных в литературе, приводит к некоторым параметрам, сомнительным с биологической точки зрения. Например, наша смертность детенышей в возрасте 0-1 года ниже, чем смертность молоди в возрасте 1-4 лет. Однако, поскольку у нас нет информации о фактическом уровне смертности бескрылых гагарок, любая корректировка этих параметров будет произвольной.Поэтому мы решили строго использовать самые низкие показатели смертности, найденные в литературе, для каждой возрастной группы. Для сравнения мы добавили симуляцию, основанную на известных показателях смертности гагарок, которые имеют более реалистичное с биологической точки зрения распределение показателей смертности, хотя, возможно, несколько слишком высокое для гагарки (см. Обсуждение и Дополнительный файл 2B)».
Кроме того, оценки возраста первого размножения и выживаемости взрослых особей, вероятно, нуждаются в корректировке.
Теперь мы сделали это в нашем анализе «реалистичных настроек» (см. Дополнительный файл 2A и B).
c) Имеется ли дополнительная историческая информация о коэффициентах вылова, такая как количество птиц или яиц, взятых из конкретной гнездовой колонии, которая позволила бы рассчитать коэффициенты вылова для конкретных мест и времени?
К сожалению, несмотря на обширные литературные поиски, нам не удалось найти такую информацию. На самом деле это было основной причиной, по которой мы решили проводить анализы с нереалистично консервативными настройками.
https://doi.org/10.7554/eLife.47509.sa2
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка браузера на прием файлов cookie
Существует множество причин, по которым файл cookie не может быть установлен правильно. Ниже приведены наиболее распространенные причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки браузера, чтобы принять файлы cookie, или спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает, хотите ли вы принимать файлы cookie, и вы отказались.
Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файл cookie. - Ваш браузер не поддерживает файлы cookie. Попробуйте другой браузер, если вы подозреваете это.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г.,
браузер автоматически забудет файл cookie. Чтобы это исправить, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.
Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Предоставить доступ без файлов cookie
потребует от сайта создания нового сеанса для каждой посещаемой вами страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в файле cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт
Например, сайт
не может определить ваше имя электронной почты, если вы не решите ввести его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступ к
остальной части вашего компьютера, и только сайт, создавший файл cookie, может его прочитать.
Вирусная геномика в исследованиях вируса Эбола
Kuhn, J.H. Filoviruses. Сборник эпидемиологических, клинических и лабораторных исследований за 40 лет. Арх. Вирол. Доп. 20 , 13–360 (2008).
Артикул
пабмед
Google Scholar
Национальный центр биотехнологической информации. GenBank https://www.ncbi.nlm.nih.gov/genbank/ (2019).
Kuhn, JH, Amarasinghe, G. & Perry, D.L. в Fields Virology : Emerging Viruses Ch. 12 (редакторы Питер Хоули, М. Найп, Дэвид М. и Уилан, Шон П.Дж.) (Wolters Kluwer, 2020).
Всемирная организация здравоохранения. Отчет о ситуации. Болезнь вируса Эбола. https://apps.who.int/iris/bitstream/handle/10665/208883/ebolasitrep_10Jun2016_eng.pdf;jsessionid=4F7DFD58B3A8FDF69F885F6DF11F4E60?sequence=1 (2016 г.).
Отчет о ситуации. Болезнь вируса Эбола. https://apps.who.int/iris/bitstream/handle/10665/208883/ebolasitrep_10Jun2016_eng.pdf;jsessionid=4F7DFD58B3A8FDF69F885F6DF11F4E60?sequence=1 (2016 г.).
Всемирная организация здравоохранения.Эбола в Демократической Республике Конго. Экстренное обновление здоровья. https://www.who.int/emergencies/diseases/ebola/drc-2019 (2020).
Whitley, R. Новый век молекулярной диагностики микробных агентов. Н. англ. Дж. Мед. 358 , 988–989 (2008).
КАС
Статья
пабмед
Google Scholar
Palacios, G. et al. Новый аренавирус в кластере фатальных заболеваний, связанных с трансплантацией. Н. англ. Дж. Мед. 358 , 991–998 (2008). Первый пример метагеномного секвенирования, демонстрирующий способность метода обнаруживать неизвестные этиологические агенты в различных типах образцов .
КАС
Статья
пабмед
Google Scholar

Liu, X. et al. Транскриптомные сигнатуры отличают выживаемость от летальных исходов у людей, инфицированных вирусом Эбола. Геном Биол. 18 , 4 (2017).
Артикул
пабмед
ПабМед Центральный
Google Scholar
McMullan, L.K. et al. Характеристика инфекционного вируса Эбола из продолжающейся вспышки для руководства ответными действиями в Демократической Республике Конго: филогенетический анализ и анализ in vitro. Ланцет Заражение. Дис. 19 , 1023–1032 (2019). Первый пример успешного использования обратной геномики филовирусов .
Артикул
пабмед
Google Scholar
Деметрия, К. и др. Повторное появление Reston ebolavirus у яванских макак, Филиппины, 2015 г. Emerg. Заразить. Дис. 24 , 1285–1291 (2018).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar

Geisbert, T.W. & Jahrling, P.B. Использование иммуноэлектронной микроскопии для выявления вируса Эбола во время эпизоотии 1989 года в США. Дж. Клин. Патол. 43 , 813–816 (1990).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Miranda, M.E.G. & Miranda, N.L.J. Reston ebolavirus у людей и животных на Филиппинах: обзор. Дж. Заражение. Дис. 204 (Приложение 3), S757–S760 (2011).
Артикул
пабмед
Google Scholar
Albariño, C.G. et al. Взгляд на побочные эффекты и адаптацию вируса Рестон на основе полных последовательностей генома вируса. PLoS One 12 , e0178224 (2017).
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar
Amman, B.R. et al. Сезонные импульсы циркуляции марбургского вируса у молоди летучих мышей Rousettus aegyptiacus совпадают с периодами повышенного риска заражения человека.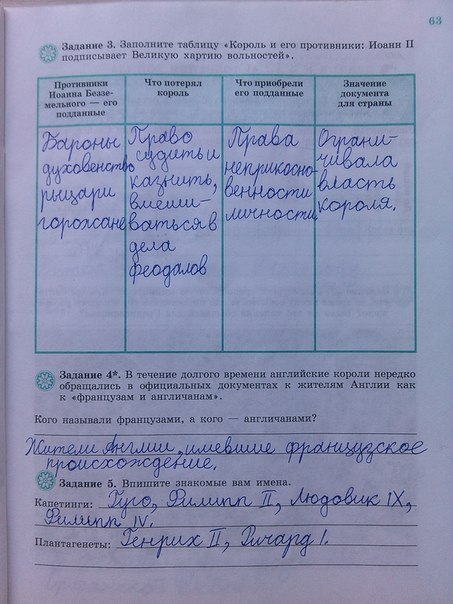 PLoS Патог. 8 , e1002877 (2012 г.).
PLoS Патог. 8 , e1002877 (2012 г.).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Павенска, Дж. Т. и др. Заражение вирусом Марбург у египетских летучих мышей, Южная Африка, 2013–2014 гг. Аварийный. Заразить. Дис. 24 , 1134–1137 (2018).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Towner, J.S. et al. Выделение генетически разнообразных вирусов Марбург от египетских летучих мышей. PLoS Патог. 5 , e1000536 (2009 г.).
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar
Амман, Б. Р. и др. Выделение анголоподобного вируса Марбург от египетских красных летучих мышей из Западной Африки. Нац. коммун. 11 , 510 (2020).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar

Swanepoel, R. et al. Исследования резервуарных хозяев марбургского вируса. Аварийный. Заразить. Дис. 13 , 1847–1851 (2007).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Каджихара, М. и др. Марбургвирус у египетских фруктовых летучих мышей, Замбия. Аварийный. Заразить. Дис. 25 , 1577–1580 (2019).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Barrette, R.W. et al. Открытие свиней как носителя эболавируса Reston . Наука 325 , 204–206 (2009).
КАС
Статья
пабмед
Google Scholar
Пан, Ю. и др. Вирус Рестона у домашних свиней в Китае. Арх. Вирол. 159 , 1129–1132 (2014).
КАС
Статья
пабмед
Google Scholar

Jayme, S. I. et al. Молекулярные доказательства заражения вирусом Эбола Рестон у филиппинских летучих мышей. Вирол. J. 12 , 107 (2015).
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar
Кеменеси, Г. и др. Повторное появление вируса Lloviu у летучих мышей Miniopterus schreibersii , Венгрия, 2016 г. Emerg. микробы заражают. 7 , 66 (2018).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Негредо, А. и др. Открытие эболавирусоподобного филовируса в Европе. PLoS Патог. 7 , e1002304 (2011). Первый филовирус, обнаруженный секвенированием нового поколения без выделения вируса .
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Forbes, K.M. et al. Вирус Bombali в Mops condylurus Bat, Кения.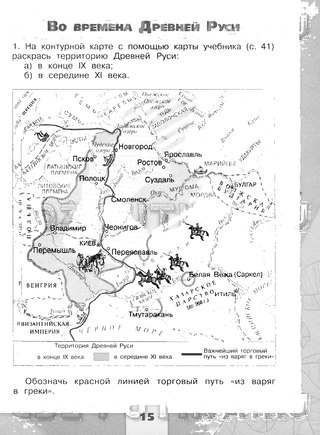 Аварийный. Заразить. Дис. 25 , 955–957 (2019).
Аварийный. Заразить. Дис. 25 , 955–957 (2019).
Артикул
ПабМед Центральный
Google Scholar
Goldstein, T. et al. Открытие вируса Бомбали подтверждает, что летучие мыши являются носителями эболавирусов. Нац. микробиол. 3 , 1084–1089 (2018).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Karan, L.S. et al. Вирус Бомбали у летучих мышей Mops condylurus , Гвинея. Аварийный. Заразить. Дис. 25 , 1774–1775 (2019).
Артикул
ПабМед Центральный
Google Scholar
Он, Б.и другие. РНК филовируса летучих мышей, Китай. Аварийный. Заразить. Дис. 21 , 1675–1677 (2015).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar

Ян, С.-Л. и другие. Характеристика филовируса (вирус Менгла) от летучих мышей Rousettus в Китае. Нац. микробиол. 4 , 390–395 (2019).
КАС
Статья
пабмед
Google Scholar
Ян, С.-Л. и другие. Генетически разнообразные филовирусы Rousettus и Eonycteris spp. летучие мыши, Китай, 2009 и 2015 гг. Emerg. Заразить. Дис. 23 , 482–486 (2017).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Shi, M. et al. Эволюционная история РНК-вирусов позвоночных. Природа 556 , 197–202 (2018). Неожиданное метагеномное открытие филовирусов у рыб .
КАС
Статья
пабмед
Google Scholar
Leroy, E.M. et al. Летучие мыши как резервуары вируса Эбола. Природа 438 , 575–576 (2005).
КАС
Статья
Google Scholar
Bermejo, M. et al. Вспышка лихорадки Эбола унесла жизни 5000 горилл. Наука 314 , 1564 (2006).
КАС
Статья
пабмед
Google Scholar
Лерой, Э. М. и др. Множественные случаи передачи вируса Эбола и быстрое сокращение дикой природы Центральной Африки. Наука 303 , 387–390 (2004).
КАС
Статья
пабмед
Google Scholar
Wittmann, T.J. et al. Изоляты эболавируса Заира от диких обезьян обнаруживают генетическое происхождение и рекомбинанты. Проц. Натл акад. науч. США 104 , 17123–17127 (2007 г.).
КАС
Статья
пабмед
Google Scholar
Холмс, Э. К., Дудас, Г., Рамбо, А. и Андерсен, К. Г. Эволюция вируса Эбола: анализ эпидемии 2013–2016 гг. Природа 538 , 193–200 (2016). В этом обзоре обобщаются усилия по секвенированию и эволюционные исследования, проведенные во время вспышки БВВЭ в Западной Африке в 2013–2016 гг. .
Природа 538 , 193–200 (2016). В этом обзоре обобщаются усилия по секвенированию и эволюционные исследования, проведенные во время вспышки БВВЭ в Западной Африке в 2013–2016 гг. .
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Baize, S. et al.Появление Заирского вируса Эбола в Гвинее. Н. англ. Дж. Мед. 371 , 1418–1425 (2014).
КАС
Статья
пабмед
Google Scholar
Gire, S.K. et al. Геномный надзор проясняет происхождение и передачу вируса Эбола во время вспышки 2014 года. Наука 345 , 1369–1372 (2014).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Ариас, А. и др. Быстрое определение последовательности вспышки вируса Эбола в Сьерра-Леоне позволяет выявить цепочки передачи, связанные со спорадическими случаями.
Артикул
пабмед
ПабМед Центральный
Google Scholar
Kugelman, J.R. et al. Мониторинг эволюции вируса Эбола Makona путем создания передовых геномных возможностей в Либерии. Аварийный. Заразить. Дис. 21 , 1135–1143 (2015). В этом отчете описывается создание первого национального центра геномики в Либерии во время вспышки БВВЭ в Западной Африке в 2013–2016 гг. .
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Quick, J. et al. Портативное секвенирование генома в режиме реального времени для эпиднадзора за лихорадкой Эбола. Природа 530 , 228–232 (2016).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Мбала-Кингебени, П. и др. Медицинские контрмеры во время вспышки болезни, вызванной вирусом Эбола, в 2018 г. в провинциях Северное Киву и Итури Демократической Республики Конго: экспресс-оценка генома. Ланцет Заражение. Дис. 19 , 648–657 (2019).
в провинциях Северное Киву и Итури Демократической Республики Конго: экспресс-оценка генома. Ланцет Заражение. Дис. 19 , 648–657 (2019).
Артикул
пабмед
Google Scholar
Mbala-Kingebeni, P. et al. Вспышка болезни, вызванной вирусом Эбола, в 2018 г. в Экваториальной провинции, Демократическая Республика Конго: ретроспективная геномная характеристика. Ланцет Заражение. Дис. 19 , 641–647 (2019).
Артикул
пабмед
Google Scholar
Mbala-Kingebeni, P. et al. Быстрое подтверждение заирского вируса Эбола во время вспышки в Экваториальной провинции Демократической Республики Конго: последствия для мероприятий общественного здравоохранения. клин. Заразить. Дис. 68 , 330–333 (2019).
Артикул
пабмед
Google Scholar
Grubaugh, N. D. et al. Отслеживание вирусных вспышек в двадцать первом веке. Нац. микробиол. 4 , 10–19 (2019).
D. et al. Отслеживание вирусных вспышек в двадцать первом веке. Нац. микробиол. 4 , 10–19 (2019).
КАС
Статья
пабмед
Google Scholar
Korber, B. et al. Время появления предка пандемических штаммов ВИЧ-1. Наука 288 , 1789–1796 (2000).
КАС
Статья
пабмед
Google Scholar
Парк, Д. Дж. и др. Эпидемиология, передача и эволюция вируса Эбола в течение семи месяцев в Сьерра-Леоне. Cell 161 , 1516–1526 (2015).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Sozhamannan, S. et al. Оценка эрозии сигнатуры вируса Эбола из-за геномного дрейфа и его влияния на эффективность диагностических тестов. Вирусы 7 , 3130–3154 (2015).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar

Blackley, D. J. et al. Снижение скорости эволюции в повторно возникших цепочках передачи вируса Эбола. науч. Доп. 2 , e1600378 (2016).
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar
де ла Вега, М.-А. и другие. Вирусная нагрузка Эболы на момент постановки диагноза связана с исходом болезни и развитием вспышки. Дж. Клин. Вкладывать деньги. 125 , 4421–4428 (2015).
Артикул
Google Scholar
Dudas, G. et al. Геномы вирусов раскрывают факторы, которые распространяли и поддерживали эпидемию лихорадки Эбола. Природа 544 , 309–315 (2017).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Мейерс Л., Фроули Т., Госс С. и Канг С. Вспышка вируса Эбола в 2014 г.: клинический обзор для врачей скорой помощи. Энн. Эмердж. Мед. 65 , 101–108 (2015).
Энн. Эмердж. Мед. 65 , 101–108 (2015).
Артикул
пабмед
Google Scholar
Urbanowicz, R. A. et al. Адаптация вируса Эбола к человеку во время вспышки в Западной Африке. Cell 167 , 1079–1087 (2016). Информация, полученная в результате секвенирования нового поколения в полевых условиях, используется для оценки влияния идентифицированных мутаций генома EBOV in vitro .
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Carroll, M.W. et al. Временной и пространственный анализ вспышки вируса Эбола в Западной Африке в 2014–2015 гг. Природа 524 , 97–101 (2015).
КАС
Статья
пабмед
Google Scholar
Nkoghé, D. et al. Cas isolé de fièvre hemorragique survenu au Gabon в 2002 году, подвеска l’épidémie d’Ebola mais remote des régions épidémiques [французский]. Мед. Троп. 65 , 349–354 (2005).
Мед. Троп. 65 , 349–354 (2005).
Google Scholar
Kerber, R. et al. Анализ результатов диагностики, полученных в европейской мобильной лаборатории в Гекеду, Гвинея, с марта 2014 г. по март 2015 г. J. Infect. Дис. 214 , С250–С257 (2016).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Лаук, М.и другие. Коинфекции вируса ГБ С у пациентов с лихорадкой Эбола в Западной Африке. Дж. Вирол. 89 , 2425–2429 (2015).
Артикул
КАС
пабмед
Google Scholar
Li, T. et al. Метагеномное секвенирование нового поколения вспышки болезни, вызванной вирусом Эбола, в 2014 году в Демократической Республике Конго. Дж. Клин. микробиол. 57 , e00827–19 (2019).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar

Kreuels, B. et al. Случай тяжелой инфекции, вызванной вирусом Эбола, осложненной грамотрицательной септицемией. Н. англ. Дж. Мед. 371 , 2394–2401 (2014).
КАС
Статья
пабмед
Google Scholar
Wolf, T. et al. Тяжелое заболевание, вызванное вирусом Эбола, с просачиванием сосудов и полиорганной недостаточностью: лечение больного в реанимации. Ланцет 385 , 1428–1435 (2015).
Артикул
пабмед
Google Scholar
Кэрролл, М. В. и др. Глубокое секвенирование РНК из образцов крови и мазков из ротовой полости выявило наличие нуклеиновых кислот ряда патогенов у пациентов с острым заболеванием, вызванным вирусом Эбола, и согласуется с бактериальной транслокацией по кишечнику. mSphere 2 , e00325–17 (2017).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar

ди Паола, Н. и др. Вспышка человеческого парвовируса B19, скрытая лихорадкой денге. клин. Заразить. Дис. 68 , 810–817 (2019).
Артикул
КАС
пабмед
Google Scholar
Delwart, E.L. Вирусная метагеномика. Rev. Med. Вирол. 17 , 115–131 (2007).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Houldcroft, C.J., Beale, M.A. & Breuer, J. Клинические и биологические данные секвенирования вирусного генома. Нац. Преподобный Микробиолог. 15 , 183–192 (2017).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Palacios, G. et al. Панмикробный олигонуклеотидный массив для диагностики инфекционных заболеваний. Аварийный. Заразить. Дис. 13 , 73–81 (2007).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar

Briese, T. et al. Секвенирование захвата вирома обеспечивает точную диагностику вируса и всесторонний анализ вирома. мБ ио 6 , e01491-15 (2015).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Chalkias, S. et al. ViroFind: новая платформа для глубокого секвенирования с обогащением мишеней выявляет сложную популяцию вируса JC в мозге пациентов с ПМЛ. PLoS One 13 , e0186945 (2018 г.).
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar
Кампершроер С., Госинк М. М., Кумпф С. В., О’Доннелл Л. М. и Тартаро К. Р. Геномная последовательность лимфокриптовируса яванского макака. Вирусология 488 , 28–36 (2016).
КАС
Статья
пабмед
Google Scholar
О’Флаэрти, Б.М. и др. Всестороннее обогащение вирусов обеспечивает идентификацию и анализ генома чувствительного респираторного вируса с помощью секвенирования нового поколения. Рез. генома. 28 , 869–877 (2018).
Рез. генома. 28 , 869–877 (2018).
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar
Kafetzopoulou, L.E. et al. Метагеномное секвенирование в эпицентре вспышки лихорадки Ласса в Нигерии в 2018 г. Наука 363 , 74–77 (2019).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Гарди, Дж. Л. и Ломан, Нью-Джерси. На пути к глобальной системе эпиднадзора за патогенами в режиме реального времени, основанной на данных геномики. Нац. Преподобный Жене. 19 , 9–20 (2018).
КАС
Статья
пабмед
Google Scholar
Ladner, J. T. et al. Эволюция и распространение вируса Эбола в Либерии, 2014-2015 гг. Микроб-хозяин клетки 18 , 659–669 (2015).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar

Mate, S.E. et al. Молекулярные доказательства передачи вируса Эбола половым путем. Н. англ. Дж. Мед. 373 , 2448–2454 (2015). Первое основанное на последовательности доказательство передачи EBOV половым путем .
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Cottam, E.M. et al. Молекулярная эпидемиология вспышки вируса ящура в Соединенном Королевстве в 2001 г. J. Virol. 80 , 11274–11282 (2006).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Worby, C.J., Lipsitch, M. & Hanage, W.P. Общие геномные варианты: идентификация путей передачи с использованием данных глубокой последовательности патогенов. утра. Дж. Эпидемиол. 186 , 1209–1216 (2017).
Артикул
пабмед
ПабМед Центральный
Google Scholar

Базелер, Л., Чертоу, Д.С., Джонсон, К.М., Фельдманн, Х. и Моренс, Д.М. Патогенез болезни, вызванной вирусом Эбола. год. Преподобный Патол. 12 , 387–418 (2017).
КАС
Статья
пабмед
Google Scholar
Мессауди, И., Амарасингхе, Г.К. и Баслер, С.Ф. Патогенез филовирусов и уклонение от иммунитета: информация о вирусе Эбола и вирусе Марбург. Нац. Преподобный Микробиолог. 13 , 663–676 (2015).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Wong, G., Kobinger, G.P. & Qiu, X. Характеристика иммунных реакций хозяина при инфекциях, вызванных вирусом Эбола. Эксперт Преподобный Клин. Иммунол. 10 , 781–790 (2014).
КАС
Статья
пабмед
Google Scholar
Матцингер, П. и Камала, Т. Контроль класса на основе тканей: другая сторона толерантности. Нац. Преподобный Иммунол. 11 , 221–230 (2011).
Нац. Преподобный Иммунол. 11 , 221–230 (2011).
КАС
Статья
пабмед
Google Scholar
Kuming, B.S. & Kokoris, N. Участие Увеала в болезни, вызванной вирусом Марбург. Бр.Дж. Офтальмол. 61 , 265–266 (1977).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Martini, G. A. & Schmidt, H. A. Spermatogene Übertragung des «Virus Marburg» (Erreger der «Marburger Affenkrankheit») [нем.]. Клин. Wochenschr. 46 , 398–400 (1968).
КАС
Статья
пабмед
Google Scholar
Rodriguez, L.L. et al. Устойчивость и генетическая стабильность вируса Эбола во время вспышки в Киквите, Демократическая Республика Конго, 1995 г. J. Infect. Дис. 179 , S170–S176 (1999).
КАС
Статья
пабмед
Google Scholar
Smith, D.H. et al. Болезнь, вызванная марбургским вирусом, в Кении. Ланцет 1 , 816–820 (1982).
КАС
Статья
пабмед
Google Scholar
Субисси, Л. и др. Передача вируса Эбола, вызванная персистентно инфицированными людьми, пережившими вспышку 2014–2016 годов в Западной Африке. Дж. Заражение. Дис. 218 , S287–S291 (2018).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Диалло, Б. и др. Возобновление болезни, вызванной вирусом Эбола, в Гвинее связано с тем, что у выжившего вирус сохраняется в семенной жидкости более 500 дней. клин.Заразить. Дис. 63 , 1353–1356 (2016).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Соу М.С. и др. Новые данные о длительной персистенции генетического материала вируса Эбола в сперме выживших.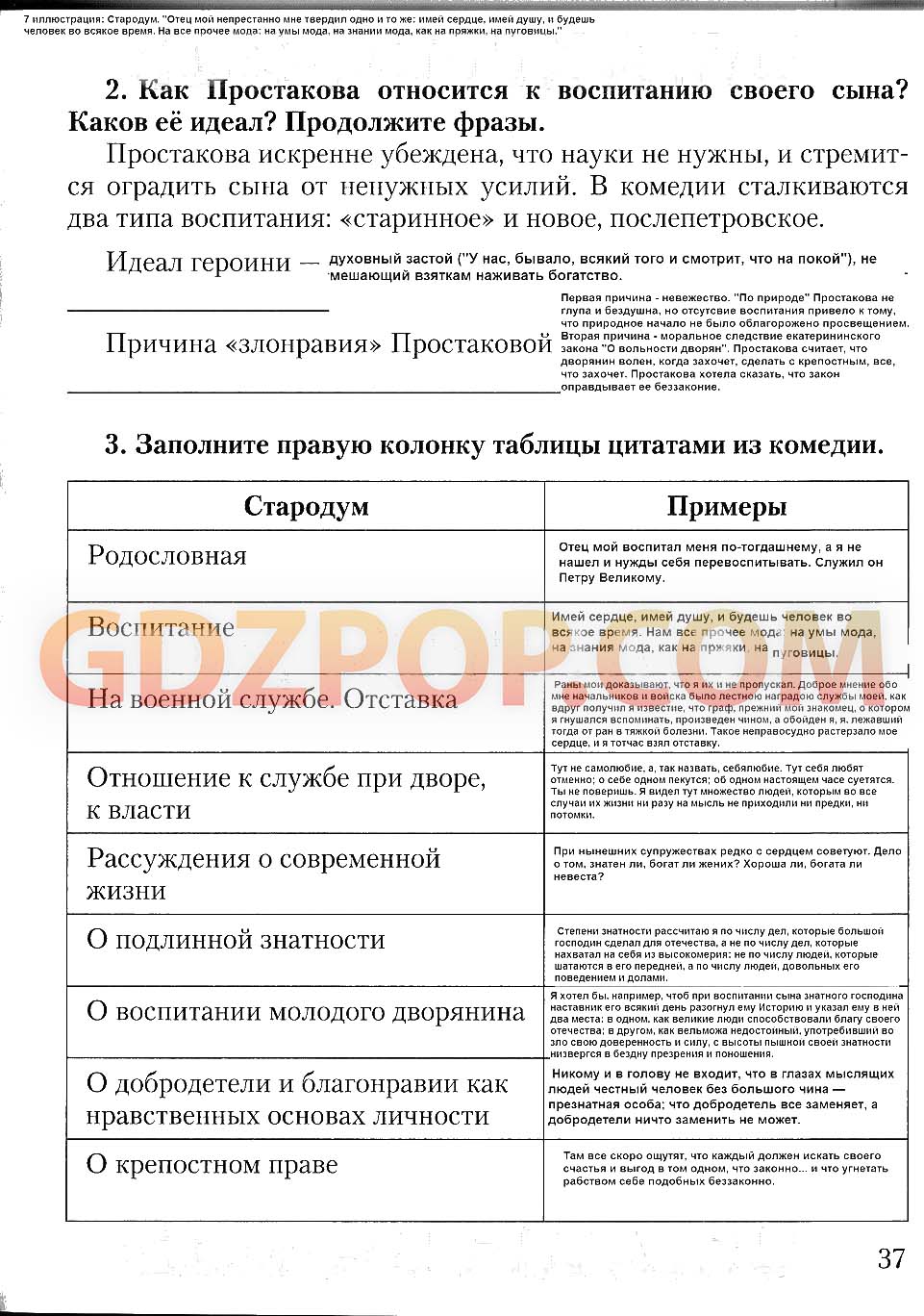 Дж. Заражение. Дис. 214 , 1475–1476 (2016).
Дж. Заражение. Дис. 214 , 1475–1476 (2016).
Артикул
пабмед
Google Scholar
Уеки Т.М. и др. Персистенция вируса Эбола в сперме выживших мужчин. клин. Заразить. Дис. 62 , 1552–1555 (2016).
Артикул
пабмед
Google Scholar
Whitmer, S.L.M. et al. Активная репликация вируса Эбола и гетерогенные скорости эволюции у выживших после БВВЭ. Cell Rep. 22 , 1159–1168 (2018).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Скотт, Дж. Т. и др. Постэбола-синдром, Сьерра-Леоне. Аварийный. Заразить. Дис. 22 , 641–646 (2016).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Веттер П., Кайзер Л., Шиблер М., Цигленецкий И. и Бауш Д. Г. Последствия болезни, вызванной вирусом Эбола: чрезвычайная ситуация внутри чрезвычайной ситуации. Ланцет Заражение. Дис. 16 , e82–e91 (2016).
и Бауш Д. Г. Последствия болезни, вызванной вирусом Эбола: чрезвычайная ситуация внутри чрезвычайной ситуации. Ланцет Заражение. Дис. 16 , e82–e91 (2016).
Артикул
пабмед
Google Scholar
Цзэн, X. и др. Идентификация и патологоанатомическая характеристика персистирующей бессимптомной вирусной инфекции Эбола у макак-резусов. Нац. микробиол. 2 , 17113 (2017).
КАС
Статья
пабмед
Google Scholar
Howlett, P. et al. Болезнь, вызванная вирусом Эбола, осложненная поздним энцефалитом и полиартритом, Сьерра-Леоне. Аварийный. Заразить. Дис. 22 , 150–152 (2016).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Jacobs, M. et al. Поздний рецидив вируса Эбола, вызывающий менингоэнцефалит: история болезни. Ланцет 388 , 498–503 (2016).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Shantha, J.G., Crozier, I. & Yeh, S. Обновленная информация о глазных осложнениях болезни, вызванной вирусом Эбола. Курс. мнение Офтальмол. 28 , 600–606 (2017).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Varkey, J.B. et al. Персистенция вируса Эбола в глазной жидкости в период выздоровления. Н. англ. Дж. Мед. 372 , 2423–2427 (2015).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Тейлор, А. В. Иммунная привилегия глаза и трансплантация. Фронт. Иммунол. 7 , 37 (2016).
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar
Deen, G.F. et al. Персистентность РНК Эбола в сперме выживших после болезни, вызванной вирусом Эбола — окончательный отчет. Н. англ. Дж. Мед. 377 , 1428–1437 (2017).
Н. англ. Дж. Мед. 377 , 1428–1437 (2017).
КАС
Статья
пабмед
Google Scholar
Роу А.К. и др. Клиническое, вирусологическое и иммунологическое наблюдение за выздоравливающими пациентами с геморрагической лихорадкой Эбола и их домашними контактами, Киквит, Демократическая Республика Конго. Дж. Заражение. Дис. 179 , S28–S35 (1999).
Артикул
пабмед
Google Scholar
Исследовательская группа PREVAIL III. Продольное исследование последствий лихорадки Эбола в Либерии. Н. англ. Дж. Мед. 380 , 924–934 (2019).
Артикул
ПабМед Центральный
Google Scholar
Schindell, B.G., Webb, A.L. & Kindrachuk, J. Персистенция и половая передача филовирусов. Вирусы 10 , 683 (2018 г.).
КАС
Статья
ПабМед Центральный
Google Scholar

Martines, R.B., Ng, D.L., Greer, P.W., Rollin, P.E. & Zaki, S.R. Тканевой и клеточный тропизм, патология и патогенез вирусов Эбола и Марбург. Дж. Патол. 235 , 153–174 (2015).
КАС
Статья
пабмед
Google Scholar
Гроб, К. М. и др. Персистирующая вирусная инфекция Марбурга в семенниках выживших нечеловеческих приматов. Клеточный микроб-хозяин 24 , 405–416.e3 (2018).
КАС
Статья
пабмед
Google Scholar
МакЭлрой, А.К. и др. Случай инфицирования человека вирусом Ласса с выраженной острой активацией Т-клеток и длительным вирус-специфическим Т-клеточным ответом. Дж. Заражение. Дис. 215 , 1862–1872 (2017).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Пшеничная Н.Ю., Сиденко И.С., Клиновая Е. П., Романова Е.Б., Журавлев А.С. Возможна передача конго-крымской геморрагической лихорадки половым путем. Междунар. Дж. Заразить. Дис. 45 , 109–111 (2016).
П., Романова Е.Б., Журавлев А.С. Возможна передача конго-крымской геморрагической лихорадки половым путем. Междунар. Дж. Заразить. Дис. 45 , 109–111 (2016).
Артикул
пабмед
Google Scholar
Richmond, J.K. & Baglole, D.J. Лихорадка Ласса: эпидемиология, клинические особенности и социальные последствия. BMJ 327 , 1271–1275 (2003).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Смит Д.Р. и др. Персистирующая инфекция вируса конго-крымской геморрагической лихорадки в семенниках и в гранулемах нечеловеческих приматов с латентным туберкулезом. PLoS Патог. 15 , e1008050 (2019).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Dokubo, E.K. et al. Сохранение вируса Эбола после прекращения широкомасштабной передачи в Либерии: отчет о вспышке. Ланцет Заражение.Дис. 18 , 1015–1024 (2018).
Ланцет Заражение.Дис. 18 , 1015–1024 (2018).
Артикул
пабмед
Google Scholar
Гупта М. и др. Персистирующая инфекция вирусом Эбола в условиях частичного иммунитета. Дж. Вирол. 78 , 958–967 (2004).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Sissoko, D. et al. Сохранение вируса Эбола в грудном молоке после незарегистрированного заболевания: вероятный источник передачи вируса от матери к ребенку. клин. Заразить. Дис. 64 , 513–516 (2017).
ПабМед
ПабМед Центральный
Google Scholar
Кэшман, К. А. и др. Иммуноопосредованный системный васкулит как предполагаемая причина внезапной сенсоневральной тугоухости после воздействия вируса Ласса на яванских макак. мБ ио 9 , e01896-18 (2018).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Liu, D. X. et al. Персистенция вируса Ласса связана с тяжелым системным артериитом у выздоравливающих морских свинок ( Cavia porcellus ). Дж. Заражение. Дис. 219 , 1818–1822 (2019).
ПабМед
Google Scholar
Becquart, P. et al. Высокая распространенность как гуморального, так и клеточного иммунитета к Заирскому эболавирусу среди сельского населения Габона. PLoS One 5 , e9126 (2010 г.).
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar
Diallo, M.S.K. et al. Распространенность инфекции среди бессимптомных и малосимптомных контактных лиц, контактировавших с вирусом Эбола в Гвинее: ретроспективное перекрестное обсервационное исследование. Ланцет Заражение. Дис. 19 , 308–316 (2019).
Дис. 19 , 308–316 (2019).
Артикул
пабмед
Google Scholar
Глинн, Дж. Р. и др. Бессимптомная инфекция и невыявленная болезнь, вызванная вирусом Эбола, в затронутых лихорадкой Эбола домохозяйствах в Сьерра-Леоне: перекрестное исследование с использованием нового неинвазивного анализа на антитела к вирусу Эбола. Ланцет Заражение. Дис. 17 , 645–653 (2017).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Leroy, E. M. et al. Бессимптомная инфекция Эбола у человека и сильная воспалительная реакция. Ланцет 355 , 2210–2215 (2000).
КАС
Статья
пабмед
Google Scholar
Moyen, N. et al. Факторы риска, связанные с серопревалентностью вирусов Эбола и Марбург у доноров крови в Республике Конго. PLoS Негл. Троп. Дис. 9 , e0003833 (2015).
Дис. 9 , e0003833 (2015).
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar
Нкоге, Д. и др. Факторы риска для Zaire ebolavirus -специфических IgG в сельском населении Габона. Дж. Заражение. Дис. 204 (Приложение 3), S768–S775 (2011).
Артикул
КАС
пабмед
Google Scholar
Ruibal, P. et al. Уникальная человеческая иммунная сигнатура болезни, вызванной вирусом Эбола, в Гвинее. Природа 533 , 100–104 (2016).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Neumann, G., Feldmann, H., Watanabe, S., Lukashevich, I. & Kawaoka, Y. Обратная генетика демонстрирует, что протеолитический процессинг гликопротеина вируса Эбола не является необходимым для репликации в клеточной культуре. Дж. Вирол. 76 , 406–410 (2002).
Вирол. 76 , 406–410 (2002).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Волчков В.Е. и др. Восстановление инфекционного вируса Эбола из комплементарной ДНК: редактирование РНК гена GP и вирусная цитотоксичность. Наука 291 , 1965–1969 (2001). Первая система обратной генетики для выделения EBOV из плазмид .
КАС
Статья
пабмед
Google Scholar
Кросс, Р. В. и Гейсберт, Т. В. Использование обратной генетики для информирования о мерах реагирования на вспышку Эболы. Ланцет Заражение. Дис. 19 , 925–927 (2019).
Артикул
пабмед
Google Scholar
Hoenen, T., Groseth, A. & Feldmann, H. Терапевтические стратегии, нацеленные на жизненный цикл вируса Эбола. Нац. Преподобный Микробиолог. 17 , 593–606 (2019).
КАС
Статья
пабмед
Google Scholar
Чисхолм, П. Дж., Буш, Дж. В. и Краудер, Д. В. Влияние истории жизни и экологии на эволюционный потенциал вируса. Вирус Рез. 265 , 1–9 (2019).
КАС
Статья
пабмед
Google Scholar
Warfield, K.L. et al. Роль антител в защите от вируса Эбола у нечеловекообразных приматов, иммунизированных тремя вакцинными платформами. Дж. Заражение. Дис. 218 , S553–S564 (2018).
Артикул
пабмед
ПабМед Центральный
Google Scholar
Wong, G. et al. Иммунные параметры коррелируют с защитой от инфекции, вызванной вирусом Эбола, у грызунов и нечеловеческих приматов. науч. Перевод мед. 4 , 158ra146 (2012).
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar

Garbutt, M. et al. Свойства репликативно-компетентных вирусных векторов везикулярного стоматита, экспрессирующих гликопротеины филовирусов и аренавирусов. Дж. Вирол. 78 , 5458–5465 (2004).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Ян, З.-Ю. и другие. Идентификация гликопротеина вируса Эбола как основной вирусной детерминанты цитотоксичности и повреждения сосудистых клеток. Нац. Мед. 6 , 886–889 (2000).
КАС
Статья
пабмед
Google Scholar
Henao-Restrepo, A.M. et al. Эффективность и действенность вакцины с вектором rVSV в профилактике болезни, вызванной вирусом Эбола: окончательные результаты вакцинации Гвинейского кольца, открытое, кластерное рандомизированное исследование (Ebola Ça Suffit!). Ланцет 389 , 505–518 (2017).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar

Национальный институт аллергии и инфекционных заболеваний. Независимый совет по мониторингу рекомендует досрочно прекратить испытания препаратов против Эболы в ДРК из-за положительных результатов двух из четырех кандидатов. https://www.niaid.nih.gov/news-events/independent-monitoring-board-recommends-early-termination-ebola-therapy-trial-drc (2019).
Mulangu, S. et al. Рандомизированное контролируемое исследование терапии болезни, вызванной вирусом Эбола. Н. англ. Дж. Мед. 381 , 2293–2303 (2019).
КАС
Статья
пабмед
Google Scholar
Corti, D. et al. Протективная монотерапия против летальной инфекции вирусом Эбола сильнодействующими нейтрализующими антителами. Наука 351 , 1339–1342 (2016).
КАС
Статья
пабмед
Google Scholar
Wec, A.Z. et al. Антитела выжившего человека определяют уязвимые места для широкой защиты от эболавирусов. Cell 169 , 878–890.e15 (2017).
Cell 169 , 878–890.e15 (2017).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Gaudinski, M.R. et al. Безопасность, переносимость, фармакокинетика и иммуногенность терапевтического моноклонального антитела mAb114, нацеленного на гликопротеин вируса Эбола (VRC 608): открытое исследование фазы 1. Ланцет 393 , 889–898 (2019).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Misasi, J. et al. Структурно-молекулярные основы нейтрализации вируса Эбола защитными человеческими антителами. Наука 351 , 1343–1346 (2016).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Паскаль, К. Э. и др. Разработка человеческих моноклональных антител на клинической стадии для лечения прогрессирующей болезни, вызванной вирусом Эбола, у нечеловекообразных приматов. Дж. Заражение. Дис. 218 (Прил. 5), S612–S626 (2018).
Дж. Заражение. Дис. 218 (Прил. 5), S612–S626 (2018).
Артикул
Google Scholar
Qiu, X. et al. Успешное лечение инфицированных вирусом Эбола яванских макак моноклональными антителами. науч. Перевод мед. 4 , 138ra81 (2012).
Артикул
пабмед
Google Scholar
Audet, J. et al. Молекулярная характеристика моноклональных антител, составляющих ZMAb: защитный коктейль против вируса Эбола. науч. 4 , 6881 (2014).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Davidson, E. et al. Механизм связывания гликопротеина вируса Эбола с помощью антител ZMapp, ZMAb и MB-003. Дж. Вирол. 89 , 10982–10992 (2015).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar

Miller, C. R. et al. Инициирование списка наблюдения за мутациями ускользания антител вируса Эбола. PeerJ 4 , e1674 (2016).
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar
Патель, Дж.С., Куэйтс, С.Дж., Джонсон, Э.Л. и Итреберг, Ф.М. Расширение контрольного списка потенциальных мутаций ускользания антител от вируса Эбола. PLoS One 14 , e0211093 (2019).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Kugelman, J.R. et al. Появление ускользающих вариантов вируса Эбола у инфицированных нечеловеческих приматов, обработанных коктейлем антител MB-003. Cell Rep. 12 , 2111–2120 (2015). Первая основанная на последовательности характеристика EBOV, избежавшего медицинских контрмер in vivo .
КАС
Статья
пабмед
Google Scholar

Johnson, W.E. Происхождение и эволюционные последствия древних эндогенных ретровирусов. Нац. Преподобный Микробиолог. 17 , 355–370 (2019).
КАС
Статья
пабмед
Google Scholar
Ми, С. и др. Синцитин представляет собой пленочный белок оболочки ретровируса, участвующий в морфогенезе плаценты человека. Природа 403 , 785–789 (2000).
КАС
Статья
пабмед
Google Scholar
Крюков К., Уэда М. Т., Иманиши Т. и Накагава С. Систематическое исследование неретровирусных вирусоподобных элементов в эукариотических геномах. Вирус Рез. 262 , 30–36 (2019).
КАС
Статья
пабмед
Google Scholar
Белый В. А., Левин А. Дж. и Скалка А. М. Неожиданное наследование: множественные интеграции последовательностей древних борнавирусов и эболавирусов/марбургвирусов в геномах позвоночных. PLoS Патог. 6 , e1001030 (2010 г.).
PLoS Патог. 6 , e1001030 (2010 г.).
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar
Horie, M. & Tomonaga, K. Палеовирусология борнавирусов: что можно узнать из молекулярных окаменелостей борнавирусов. Вирус Рез. 262 , 2–9 (2019).
КАС
Статья
пабмед
Google Scholar
Chiba, S. et al. Широко распространенная эндогенизация геномных последовательностей неретровирусных РНК-вирусов в геномы растений. PLoS Патог. 7 , e1002146 (2011).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
ter Horst, A.M., Nigg, JC, Dekker, F.M. & Falk, B.W. Эндогенные вирусные элементы широко распространены в геномах членистоногих и обычно дают начало взаимодействующим с PIWI РНК. Дж. Вирол. 93 , e02124–18 (2019).
ПабМед
ПабМед Центральный
Google Scholar
Taylor, D.J., Dittmar, K., Ballinger, M.J. & Bruenn, J.A. Эволюционное сохранение филовирусоподобных генов в геномах летучих мышей. BMC Evol.биол. 11 , 336 (2011).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
Taylor, D.J., Leach, R.W. & Bruenn, J. Филовирусы являются древними и интегрированы в геномы млекопитающих. BMC Evol. биол. 10 , 193 (2010). Основанные на последовательности доказательства долгой совместной эволюции филовирусов и филовирусов-хозяев млекопитающих .
Артикул
КАС
пабмед
ПабМед Центральный
Google Scholar
Gu, W., Miller, S. & Chiu, C. Y. Клиническое метагеномное секвенирование следующего поколения для обнаружения патогенов. год. Преподобный Патол. 14 , 319–338 (2019).
Преподобный Патол. 14 , 319–338 (2019).
КАС
Статья
пабмед
Google Scholar
Faria, N.R. et al. Создание и скрытая передача вируса Зика в Бразилии и Америке. Природа 546 , 406–410 (2017).
КАС
Статья
пабмед
ПабМед Центральный
Google Scholar
404 Страница ошибки
ISSN-1550-7521
Индекс h5 Google Scholar: 7
Импакт-фактор Research Gate: 0.27
Значение SJR-0,11, индекс H-6
Global Media Journal предлагает платформу с открытым доступом для ученых, исследователей, академиков и студентов, изучающих СМИ и коммуникации, с целью публикации самых актуальных и инновационных исследований, изучающих СМИ, общество и культуру в свете глобализации.
В сочетании с революцией в области информационных и коммуникационных технологий глобализация может привести к феноменальным изменениям в области массовой коммуникации во всем мире.Журнал стремится фиксировать основные события в медиа-индустрии, которая растет вертикально и горизонтально. Влияние глобальных медиа в обществах без границ столь же огромно. Журнал поощряет оригинальные исследования СМИ об обществе и культурах, затрагивающие экономику класса, пола, возраста, региона и религии.
Заявление об открытом доступе
GMJ соответствует требованиям Будапештской инициативы по открытому доступу (BOAI), Берлинской декларации об открытом доступе к знаниям в области естественных и гуманитарных наук, Заявлению Bethesda о публикациях в открытом доступе.Окончательная опубликованная онлайн-версия статьи становится немедленно доступной для загрузки, повторного использования и распространения по всему миру с надлежащим указанием авторства. Помимо архива журнала, статья может быть размещена автором или издателем в любом электронном репозитории открытого доступа.
Turkceingilizce.gen.tr ile turkce ingilizce ceviri, cumle ceviri, dil ceviri ya da ingilizce turkce ceviri hizmetlerinden ucretsiz yararlanabilirsiniz.
ingilizceturkce.gen.tr ingilizce turkce ceviri, turkce ingilizce ceviri, ingilizce turkce cumle ceviri, ingilizceyi turkceye ceviri ve turkceyi ingilizceye ceviri gibi tum dillerde dogru ceviri hizmetleri sunuyor.
После двойного рецензирования журнал приглашает рукописи в качестве исследовательских статей, обзоров, тематических исследований, комментариев и авторских статей в таких областях, как политическая экономия средств массовой информации с подробным описанием прав собственности и контроля, телевизионных жанров глобальной эпохи, появления средств массовой информации. конгломерации, проблемы и вызовы, связанные с управлением, медиа как культурная индустрия, культурные практики, основанные на потреблении и образе жизни, регулирование и контроль, этика СМИ в условиях конкуренции в преимущественно капиталистических условиях, коммерциализация новостей, новые медиатехнологии, глобальные vs. местные, альтернативные медиа и множество проблем, которые могут возникать время от времени.
местные, альтернативные медиа и множество проблем, которые могут возникать время от времени.
Global Media Journal играет потенциальную роль в областях политической экономии средств массовой информации, подробно описывая право собственности и контроль, телевизионные жанры глобальной эпохи, появление медиаконгломератов, вопросы и проблемы, связанные с управлением, СМИ как культурная индустрия, культурные практики. сосредоточены на потреблении и образе жизни, регулировании и контроле, этике СМИ в связи с конкуренцией в преимущественно капиталистических условиях, коммерциализации новостей, новых медиатехнологиях, глобальном и глобальном.местные, альтернативные СМИ и множество проблем, которые могут возникать время от времени.
Уважая разнообразие и мультикультурализм, журнал продвигает междисциплинарные исследования в области медиа-исследований. Журнал удовлетворяет потребности широкого круга аудитории, включая студентов, хорошо разбирающихся академиков, сезонных исследователей и новаторов в области медиа и массовых коммуникаций, а также передовые институты, заинтересованные в продвижении сценаристов нового поколения, продюсеров и предпринимателей.
Журнал Global Media затрагивает разнообразные интересы студентов, преподавателей, ученых, исследователей и организаций, занимающихся международной деятельностью, в частности.
С момента своего открытия Global Media Journal был доступен для заинтересованных лиц бесплатно (открытый доступ). Этот журнал обеспечивает немедленный открытый доступ к своему содержанию, исходя из того принципа, что свободный доступ к исследованиям для общественности способствует более широкому глобальному обмену знаниями.
Обратите внимание, что GMJ находится на платформе открытого доступа, поэтому плата за обработку и публикацию будет взиматься со всех представленных статей после их принятия.
Canadian Science Publishing
Коммуникация, образование, история и философия
Vol. 92№. 11 стр. 1441–1445
В данной работе исследуется зависящее от времени дренирование тонкой пленки жидкости Эйринга переменной толщины, стекающей по вертикальной пластине. Получены точные решения для распределения скорости, расхода, средней толщины, средней скорости, вектора завихренности и напряжения. Представлены и обсуждены графические результаты для различных переменных параметров потока, таких как профиль скорости и переменная толщина.
Получены точные решения для распределения скорости, расхода, средней толщины, средней скорости, вектора завихренности и напряжения. Представлены и обсуждены графические результаты для различных переменных параметров потока, таких как профиль скорости и переменная толщина.
Исследования в области образования по физике
Vol. 92№. 11 стр. 1297–1300
Определяют распределение электрического потенциала в очень длинной пластине (например, длинной металлической линейке). Это достигается конформным отображением пластины на плоскость, что упрощает геометрию граничных условий. Обсуждаются особенности потенциала, а также их регуляризация конечным размером электрических контактов. Указывается аналогия с перенормировкой. Результаты сравниваются с предыдущими исследованиями.
Математическая физика
Том. 92№. 11 стр. 1320–1323
В этой статье мы рассматриваем вращающуюся заряженную черную дыру в трех измерениях со скалярным зарядом и изучаем некоторые гидродинамические величины, такие как константа диффузии, сдвиговая вязкость и проводимость. Мы изучаем влияние параметров черной дыры на гидродинамические величины. Мы подтвердили нижнюю границу отношения сдвиговой вязкости к энтропии.
Мы изучаем влияние параметров черной дыры на гидродинамические величины. Мы подтвердили нижнюю границу отношения сдвиговой вязкости к энтропии.
Том. 92№. 11 стр. 1337–1349
Эта статья посвящена проблеме полной периодической антисинхронизации нейронных сетей на основе мемристоров со смешанными изменяющимися во времени задержками.В рамках решений Филиппова дифференциальных уравнений с разрывной правой частью, основанных на теореме Мавина о совпадениях в теории многозначного анализа, представлено доказательство существования периодического решения. С помощью функционального подхода Ляпунова–Красовского конструируется адаптивный регулятор и неизвестные параметры управления ведомой системы определяются адаптивными законами, а также решается условие полной периодической адаптивной антисинхронизации для обеспечения глобальной антисинхронизации ведомой системы с ведущей системой.Приведен наглядный пример, демонстрирующий эффективность полученных результатов.
Том. 92№. 11 стр. 1380–1386
11 стр. 1380–1386
Мы показываем, что при выводе частоты прецессии Томаса учитывается факт осуществления безвращательных преобразований Лоренца между лабораторной системой отсчета K L и системами Лоренца K( t ), движущимися совместно с частицей со спином в любые моменты времени, t , имеет принципиальное значение. Выбирая для наблюдения за движением частицы любую другую инерциальную систему отсчета K, связанную с K L преобразованием без вращения, мы должны понимать, что преобразования между K и K( t ) при любых t равны больше не без вращения.Таким образом, мы даем разрешение известного парадокса Бакри (H. Bacry. Nuovo Cimento, 26, 1164 (1962)) и предлагаем новую интерпретацию прецессии Томаса, которая обсуждается далее.
Том. 92№. 11 стр. 1397–1399
Проведенные ранее сравнения кривых вращения галактик с MOND привели к выводу, что параметр a 0 лежит в пределах ∼20% от скорость света, а H 0 — постоянная Хаббла. Здесь предполагается, что при этом значении H 0 сигналы, распространяющиеся по периферии Вселенной, синхронизируются по фазе гравитон-нуклонным взаимодействием.
Здесь предполагается, что при этом значении H 0 сигналы, распространяющиеся по периферии Вселенной, синхронизируются по фазе гравитон-нуклонным взаимодействием.
Том. 92№. 11 стр. 1474–1480
Целью этой работы является изучение гравитационного коллапса в пространстве-времени Вайдья, встроенного в теорию гравитации Галилеона. Галилеонная гравитация на самом деле является инфракрасной модификацией гравитации Эйнштейна, которая была предложена как обобщение четырехмерной эффективной теории в модели бран DGP.Метрика Вайдьи используется повсюду, чтобы проследить характер будущих исходящих радиальных нулевых геодезических. Основной целью является определение того, является ли центральная сингулярность голой или покрытой горизонтом событий, по наличию направленной в будущее радиальной нулевой геодезической, испускаемой в прошлом из сингулярности. Чтобы указать на существование положительного касательного решения траектории, были применены как частные параметрические случаи (через табличные формы), так и широкодиапазонный процесс оконтуривания. Именно уравнение состояния (EoS) в идеальной жидкости удовлетворяет широкому спектру явлений: от пыли до экзотических жидкостей, таких как темная энергия.Мы использовали параметр EoS, k , для определения конечного состояния коллапса в разные космологические эпохи.
Именно уравнение состояния (EoS) в идеальной жидкости удовлетворяет широкому спектру явлений: от пыли до экзотических жидкостей, таких как темная энергия.Мы использовали параметр EoS, k , для определения конечного состояния коллапса в разные космологические эпохи.
Том. 92№. 11 стр. 1481–1484
В этой статье мы рассматриваем заряженную черную дыру в трех измерениях со скалярным зарядом и обсуждаем потери энергии тяжелой частицы, движущейся вблизи горизонта черной дыры. Этот анализ полезен, когда применяется соответствие антипространства де Ситтера и конформной теории поля. Мы находим, что электрический заряд черной дыры увеличивает силу сопротивления, а скалярный заряд уменьшает ее.
Том. 92№. 11 стр. 1485–1488
В этой статье мы обнаруживаем, что уравнения для гравитации могут быть адаптированы путем определения уравнений кривизны пространства-времени в терминах геодезических. Используя эти уравнения, мы переводим эту кривизну обратно в уравнения для расширенной ньютоновской силы гравитации. Используя проработанные примеры, мы можем показать, что усовершенствованные уравнения Ньютона дают результаты, технически точно согласующиеся с гравитационным экспериментом. Эти уравнения также технически точно согласуются с данными о двойных пульсарах.В то же время эти гравитационные уравнения разрешают трудности с образованием сингулярностей. Важно отметить, что передовая ньютоновская гравитация обеспечивает легко проверяемые гравитационные предсказания, особенно вблизи черных дыр.
Используя проработанные примеры, мы можем показать, что усовершенствованные уравнения Ньютона дают результаты, технически точно согласующиеся с гравитационным экспериментом. Эти уравнения также технически точно согласуются с данными о двойных пульсарах.В то же время эти гравитационные уравнения разрешают трудности с образованием сингулярностей. Важно отметить, что передовая ньютоновская гравитация обеспечивает легко проверяемые гравитационные предсказания, особенно вблизи черных дыр.
Элементарные частицы и ядерная физика
Том. 92№. 11 стр. 1400–1404
В этой статье эксклюзивный распад B+→Ds*+ ϕ рассчитывается с использованием метода факторизации КХД (QCDF) и взаимодействия в конечном состоянии (FSI). Во-первых, распад B+→Ds*+ ϕ рассчитывается методом QCDF, и в этом методе существуют только графы аннигиляции.Результат, полученный с помощью метода QCDF, ниже экспериментального результата. Считается, что FSI решает распад B+→Ds*+ ϕ. Для этого распада в качестве промежуточных состояний выбраны D + K 0 , D 0 K + и Ds+j/ψ через обмен K 0 , K + и Ds+ и вычисляем B + → D + K 0 → распад Ds* +ϕ. Амплитуда распада B + → D + K 0 снова рассчитывается методом QCDF.Экспериментальный коэффициент ветвления распада B+→Ds*+ ϕ составляет менее 1,2 × 10 –5 , а наши результаты, рассчитанные с использованием метода QCDF и FSI, составляют (0,4 ± 0,06) × 10 –7 и (0,93 ± 0,08) × 10 −5 соответственно.
Том. 92№. 11 стр. 1411-1418
Взаимодействие 48 Ca сферическое ядро рассматриваются с деформированными мишенями 224 RA и 244 PU на образование супер тяжелых элементов 272 HS и 292 114 ( 292 FL) , соответственно.Модель двойной укладки с эффективной силой M3Y-NN, зависящей от плотности, и метод функционала плотности энергии, основанный на силе Скирма, используются для получения взаимодействия ядро-ядро. Изучается влияние деформации и ориентации на параметры кулоновского барьера, и результаты сравниваются с соответствующими величинами, полученными из простой модели, основанной на приближенном подходе для ядерной части и простой аналитической формуле для кулоновского взаимодействия. Согласованное поведение результатов достигается в определенных диапазонах параметров деформации и ориентации.
Том. 92№. 11 стр. 1453–1459
Методом эйконала построены дифференциальные уравнения для пучков электромагнитных волн в магнитном и гравитационном полях пульсаров, согласно нелинейной электродинамике вакуума. Интегрирование этих уравнений с помощью последовательных приближений позволяет найти уравнения лучей и параметрическую форму закона движения электромагнитных импульсов этих лучей. Показано, что электромагнитный импульс, проходя через магнитное поле пульсара, распространяется как две нормальные волны, приходя к детектору по двум разным лучам и проводя в пути разное время.Рассчитаны углы искривления лучей из-за нелинейности электродинамики вакуума и время задержки электромагнитного сигнала, переносимого одной нормальной волной относительно другой нормальной волны. Оценивается величина этих эффектов.
Атомная и молекулярная физика
Том. 92№. 11 стр. 1285–1296
Мы представляем точные расчеты атомной структуры для 200 нижних уровней тонкой структурной энергии кислородоподобного никеля, который может быть полезным ионом как для астрофизической, так и для термоядерной плазмы.Для расчетов уровней энергии и интенсивности излучения мы использовали многоконфигурационный метод Дирака–Фока. Наши результаты сравниваются с результатами, полученными с использованием других численных методов и экспериментов, чтобы можно было оценить их точность. Приведены длины волн переходов, силы осцилляторов и скорости излучения для электрических дипольных (E1) переходов из основного состояния. Мы также представили вероятности перехода и длину волны некоторых запрещенных переходов. Наконец, мы предсказываем новые уровни энергии, силы осцилляторов и данные о вероятности перехода, когда нет никаких других теоретических или экспериментальных результатов, которые могут быть полезны для будущей экспериментальной работы.
Том. 92№. 11 стр. 1301–1304
Рентгеновские фотоны K-оболочки Fe, Ag и Te после распада электронного захвата Co 57 , Cd 109 и I 125 соответственно были обнаружены с использованием Si (Li) детектор, соединенный с многоканальным анализатором 8k на базе ПК с использованием простого метода, предложенного ранее нашей группой. Рассчитанные отношения интенсивности рентгеновского излучения K-оболочки и вероятности переноса вакансии для этих элементов сравниваются с теоретическими значениями, предсказанными Скофилдом, и с другими экспериментальными значениями, полученными с помощью фотоионизации.Установлено, что отношение интенсивностей рентгеновского излучения Κ β к Κ α Fe меньше на 4,0 %, а Ag и Te больше на 7,9 % и 19,1 % соответственно от теоретических значений. Эти отклонения могут быть связаны с обменными взаимодействиями между электронами 3p–3d-оболочек, а также с эффектом отдачи ядра за счет испускания нейтрино.
Том. 92№. 11 стр. 1330–1336
В возбужденном состоянии ( μ e ) дипольный момент молекулы является важным параметром, дающим информацию об электронной и геометрической структуре молекулы в короткоживущем состоянии.Мы экспериментально оценили дипольный момент в возбужденном состоянии ( μ e ) двух новых производных кумарина, а именно диэтил 2-ацетамидо-2-((6-метил-2-оксо-2H-хромен-4-ил) метил) маламут (DAM) и 5,6-бензо-3-[1-(4,5-дикарбометокси-1,2,3-триазолоацетил)] кумарин (5BDTC) тремя методами: Липперта, Бахшиева и Кавского. – Чамма – Виалле. В методе сольватохромного сдвига спектральный сдвиг обусловлен внутренним возбуждением молекулы. Нами были зарегистрированы спектры поглощения и испускания двух указанных выше новых производных кумарина при комнатной температуре с использованием растворителей различной полярности.Геометрия молекул была полностью оптимизирована, а дипольный момент основного состояния ( μ g ) также был рассчитан теоретически с помощью программного обеспечения Gaussian 03 с использованием уровня теории B3LYP/6-31g*. Дипольный момент в возбужденном состоянии рассчитывали на уровне теории CIS/6-31G*. µ g и µ e были рассчитаны с использованием метода сольватохромного сдвига, а µ e также были определены в сочетании с µ g 9013.Было замечено, что μ e были больше, чем μ g , что указывает на существенное перераспределение π -электронных плотностей в более полярном возбужденном состоянии для этого кумарина. Далее рассчитывали изменения дипольного момента (Δ μ ) как по методу сольватохромного сдвига, так и на основе микроскопической эмпирической полярности растворителя (ETN) и сравнивали полученные значения.
Том. 92№. 11 стр. 1405–1410
В нашей предыдущей статье ( Can.Дж. Физ. 91, 715 (2013) doi: 10.1139/cjp-2013-0077) изучалась система, состоящая из протона, мюона и электрона; мюон и электрон находятся в круговых состояниях. Исследование было мотивировано многочисленными применениями мюонных атомов и молекул, где один из электронов заменен более тяжелым лептоном μ – . Мы продемонстрировали, что в такой квазимолекуле μpe мюонное движение может представлять собой быструю подсистему, в то время как электронное движение может представлять собой медленную подсистему — результат, который может показаться нелогичным.Другими словами, мюон быстро вращается по круговой орбите вокруг оси, соединяющей протон и электрон, в то время как эта ось медленно вращается вслед за относительно медленным движением электрона. Мы показали, что спектральные линии, излучаемые мюоном в квазимолекуле μpe, испытывают красное смещение по сравнению с соответствующими спектральными линиями, которые были бы излучены мюоном в мюонном атоме водорода. В настоящей работе мы обобщаем это исследование, заменяя протон в квазимолекуле μpe полностью лишенным атома с ядерным зарядом Z > 1.Мы показываем, что в этом случае, как и в ранее изученном случае Z = 1, мюонное движение может представлять собой быструю подсистему, а электронное движение может представлять собой медленную подсистему. Для этого отношение мюонного и электронного угловых моментов должно быть несколько больше, чем в случае Z = 1. Мы показываем, что энергии связи мюона для Z > 1 значительно больше, чем для Z > 1. Z = 1 при любом конечном значении расстояния ядро–электрон.Наконец, мы показываем, что красное смещение спектральных линий, испускаемых мюоном (по сравнению со спектральными линиями соответствующего мюонного водородоподобного иона с ядерным зарядом Z ), уменьшается по мере увеличения Z . Однако относительное красное смещение остается в пределах спектрального разрешения доступных спектрометров, по крайней мере, до Z = 5. Наблюдение этого красного смещения должно быть одним из способов обнаружения образования квазимолекул μ Z e.
Том. 92№.11 стр. 1425–1429
Вероятности электродипольных переходов и силы осцилляторов нейтрального скандия рассчитаны с использованием теории орбиталей квантовых дефектов и кода Коуэна для 3d4s , и 3d4s 2 –4s 2 переходы 4p. Кроме того, с использованием кода Коуэна выполнен параметрический анализ тонкой структуры для конфигурации 3d4s 2 четной четности. Вероятности перехода и силы осцилляторов, полученные в этой работе, в основном согласуются с данными, полученными из различных теоретических работ, приведенных в литературе.Кроме того, впервые приведены вероятности перехода для 24 уровней нейтрального скандия.
Том. 92№. 11 стр. 1489–1493
K Оболочечные рентгеновские лучи бария и таллия после распада внутренней конверсии в Cs 137 и Hg 203 соответственно, были обнаружены с использованием Si(Li) рентгеновского детектора, соединенного с многоканальным 8k на базе ПК анализатор, использующий метод, предложенный ранее нашей группой. Рассчитаны отношения интенсивностей рентгеновского излучения K-оболочки и вероятности переноса вакансии для таллия и бария.Полученные результаты сравниваются с теоретическими, полуэмпирическими и другими экспериментальными результатами, полученными с помощью процессов фотоионизации, а также процессов распада. Обсуждается влияние бета-распада и внутренней конверсии на вероятности рентгеновского излучения.
Газы, гидродинамика и плазма
Vol. 92№. 11 стр. 1305–1311
Исследуется двумерная задача в трансверсально-изотропной термоупругой среде без диссипации энергии и с двумя температурами, обусловленными тепловым источником.В качестве приложения проблемы был взят конкретный тип непрерывного источника тепла, чтобы проиллюстрировать полезность подхода. Задача решается численно с использованием метода конечных элементов. Компоненты смещения, кондуктивная температура и компоненты напряжения были получены численно и проиллюстрированы графически для нашей конкретной модели. Обсуждаются также некоторые частные случаи, представляющие интерес. Для реализации кодов метода конечных элементов использовался C++. Численная работа также выполняется для подходящего материала с целью иллюстрации результатов.
Том. 92№. 11 стр. 1312–1319
Одним из сложных моментов при моделировании течения наножидкости через пористую среду является моделирование поверхностного теплового потока в присутствии наночастиц и внутренней твердой матрицы. Вопрос в том, сколько энергии поглощается твердой фазой, жидкой фазой и частицами на поверхности наложенного теплового потока? Чтобы получить подходящий ответ, в этой статье представлен локальный тепловой неравновесный подход (включая три уравнения энергии) и впервые предложены три модели теплового потока.Предложенные модели сравниваются и анализируются. Полностью изучено влияние межузельных коэффициентов теплоотдачи на теплообмен в пористом канале. Полностью изучены и сопоставлены распределения температуры жидкости и скорости теплообмена, полученные гомогенным и неоднородным подходами (для предложенных моделей). Результаты показывают, что неоднородный подход имеет большее число Нуссельта, чем однородный, для всех рекомендуемых моделей теплового потока.
Том.92№. 11 стр. 1387–1396
В этом исследовании исследуются эффекты вязкой и омической диссипации при стационарной, ламинарной, смешанной, конвекционной теплопередаче для электропроводящей жидкости, протекающей по вертикальному каналу, как в условиях поддержки, так и в условиях противодействия плавучести. Пластины обмениваются теплом с внешней жидкостью. Рассматриваются как условия равных, так и различных эталонных температур внешней жидкости. Во-первых, более простые случаи пренебрежимо малого числа Бринкмана или пренебрежимо малого числа Грасгофа рассматриваются с помощью аналитических решений.Совместное влияние сил плавучести и вязкой диссипации анализируется с использованием метода рядов возмущений, справедливого для малых значений параметра возмущения. Чтобы ослабить условия на параметр возмущения, основные уравнения также оцениваются численно с помощью метода стрельбы, который использует классический явный метод четырех наклонов Рунге-Кутты в качестве схемы интегрирования и метод Ньютона-Рафсона в качестве схемы коррекции. В рассмотренных случаях полей скорости и температуры исследуются числа Нуссельта как на стенках, так и средняя скорость.Установлено, что профили скорости для разомкнутой цепи ( E > 0 или E < 0) лежат между коротким замыканием ( E = 0). Представлены графические результаты, иллюстрирующие влияние различных параметров на поток, а также среднюю скорость и числа Нуссельта для разомкнутых и коротких цепей. В отсутствие параметра нагрузки электрического поля и числа Гартмана результаты согласуются с Zanchini ( Int. J. Heat Mass Transfer , 41, 3949 (1998)).Далее, аналитическое и численное решения очень хорошо согласуются при малых значениях параметра возмущения.
Том. 92№. 11 стр. 1419–1424
Теоретически предложен линейный анализ устойчивости простой политропической модели динамики солнечного ветра в рамках магнитогидродинамической равновесной конфигурации. Упрощенный анализ основан на модели, разработанной на основе данных, полученных в результате миссии космического корабля Advanced Composition Explorer (ACE). Уникальная форма дисперсионного уравнения получена путем объединения адиабатических и политропных процессов в пределе приближения идеального газа для газа солнечного ветра в соответствии со стандартным методом Фурье.Применяя обычную методологию разделения переменных к дисперсионному уравнению, мы получаем линейную скорость роста флуктуаций. Видно, что скорость роста является явно нелинейной функцией переменного индекса политропы ( α ) и радиального положения ( r ) относительно рассматриваемого центра Солнца. Численный анализ проводится для понимания физического понимания профилей роста флуктуаций. Показано, что рост максимален вблизи солнечной короны, где α ∼ 1, по сравнению с наблюдаемым в других местах во всей солнечной плазменной системе.Источником этого роста может быть свободный поток энергии, исходящий от динамического равновесия самой солнечной плазмы. По сравнению с существующими модельными предсказаниями наши результаты качественно способны воспроизвести среднее поведение флуктуаций и устойчивости солнечного ветра в астрофизических масштабах пространства и времени.
Конденсированные вещества
Том. 92№. 11 стр. 1356–1358
Рассмотрены длинноволновые фонон-поляритоны и продольные оптические фононы в щелочно-галоидных ионных кристаллах.Используется модель точечных зарядов, поляризованных в самосогласованном электромагнитном поле в диэлектрической среде. Получены стандартные законы дисперсии как для ветвей фонон-поляритонов, так и для продольных оптических фононов. Частота поперечного оптического фонона находится из условия электростатического равновесия. Сравнением с табличными данными доказано, что найденная частота совпадает с плазменной частотой ионов, умноженной на соотношение (ε∞+2)/[ 3(ε0−ε∞) ], где ε ∞ и ε 0 – высокочастотная диэлектрическая и статическая постоянные соответственно.
Том. 92№. 11 стр. 1359–1371
Исследование посвящено вращению магнитотермоупругого твердого тела с пустотами, подверженного тепловому нагружению лазерным импульсом. Ограничивающая плоская поверхность нагревается негауссовым лазерным лучом. Вся пористая среда вращается с равномерной угловой скоростью. Задача исследуется в рамках теории Грина-Нагди (ГН) типов II и III с учетом вращения, магнитного поля, тепловой нагрузки и пустот. Анализ нормальных мод используется для решения физической задачи для получения точных выражений для компонент перемещений, напряжений, распределения температуры и изменения поля объемных долей, которые показаны графически путем сравнения двух типов теории ГН (типы II и III) при наличии и отсутствии вращения и магнитного поля и для двух значений времени на термоупругом материале с пустотами.
Том. 92№. 11 стр. 1372–1379
Процессы рассеяния электронов на короткодействующем потенциале, обусловленные взаимодействием с полярными и неполярными оптическими фононами, пьезоэлектрическими и акустическими фононами, статической деформацией, нейтральными и ионизированными примесями в вюртците n-ZnO с концентрацией примеси ∼1 × 10 17 см −3 . Рассчитаны температурные зависимости подвижности электронов и фактора Холла в диапазоне 15/550 К.
Том. 92№. 11 стр.1446–1452
Икосаэдрическая симметрия широко распространена в природе, и понимание возможных деформаций структур, демонстрирующих ее, может иметь решающее значение для определения фундаментальных свойств. В этой работе мы представляем основу для создания и представления деформаций таких структур при сохранении икосаэдрической симметрии. Это делается путем рассмотрения точек орбиты группы икосаэдра как вершин икосаэдрического многогранника. Сжатие орбиты определяется как непрерывное изменение координат доминирующей точки, задающей орбиту в соответствующем базисе, в сторону меньших положительных значений.Точная икосаэдрическая симметрия сохраняется на любой стадии сжатия. Все икосаэдрические орбиты или многогранники могут быть построены последовательными сокращениями. Это определение сжатия является общим и может быть применено к орбитам любой конечной группы отражений.
Том. 92№. 11 стр. 1464–1469
Расчеты из первых принципов были выполнены для исследования структурной устойчивости, упругих, решеточных динамических и термодинамических свойств Na 2 Se при высоком давлении. Наши результаты показывают, что Na 2 Se в фазе антифлюоритной структуры сохраняет динамическую стабильность до 30 ГПа.Также рассчитываются и обсуждаются упругие константы и термодинамические величины при высоком давлении.
Междисциплинарная, классическая и прикладная физика
Vol. 92№. 11 стр. 1324–1329
Изучается эволюция звука в среде, свойства которой необратимо изменяются в процессе распространения волны. Например, рассматривается газ, являющийся частным случаем ньютоновской жидкости. Построены кривые гистерезиса, графические изображения необратимого затухания звуковой энергии, в плоскости термодинамических состояний.Необратимые потери внутренней энергии пропорциональны полному затуханию и зависят от интенсивности и формы звуковой волны. Обсуждаются и сравниваются кривые и петли для некоторых периодических (включая пилообразную волну) и апериодических импульсных звуков.
Том. 92№. 11 стр. 1350–1355
В настоящей работе теоретически исследовано влияние малой толщины CdTe на оптические, фронтальные и обратные рекомбинационные потери и, следовательно, на эффективность солнечного элемента CdS/CdTe. Установлено, что оптические потери составляют около 23–24 % и слабо зависят от толщины CdTe.Рекомбинационные потери составляют около 28% при d CdTe = 0,45 мкм и уменьшаются до 23% при d CdTe = 1,1 мкм, поскольку рекомбинационные потери оказывают значительное влияние на более тонкие слои. Зарегистрированная эффективность находится в диапазоне 9,5–10,5 %, что соответствует толщине CdTe 0,45–1,1 мкм, и считается, что она хорошо согласуется с экспериментальными результатами. Длина диффузии электронов находится в диапазоне 1,6-15,8 мкм и соответствует 10 -9 -10 -7 с времени жизни электрона, достаточного для того, чтобы плотность тока достигла своего максимального значения (16.3 мА/см 2 ) при d CdTe = 1,1 мкм с эффективностью 10,5%. расчетная плотность тока составляет около 20 мА/см 2 и соответствующий КПД составляет 13%. Настоящие результаты приводят к сокращению разрыва между теоретическими и практическими результатами и способствуют повышению эффективности элементов CdS/CdTe в будущем.
Том.92№. 11 стр. 1430–1440
В этой статье мы описываем магнитную систему с α-винтовым вырезом и катушкой, предназначенную для быстрого переключения магнитного поля. Была предпринята попытка определить, в какой степени такая конфигурация магнита может быть эффективно использована для приборов ядерного магнитного резонанса (ЯМР) с быстрым циклом поля (FFC). В дополнение к типичным техническим требованиям (высокое отношение напряженности поля к мощности, достаточные электрические характеристики для быстрого переключения магнитного поля и однородность магнитного поля, совместимая с ЯМР), настраиваемая однородность в объеме образца и более равномерное рассеивание тепла вдоль корпус магнита входит в комплект.Оказалось, что этим требованиям очень хорошо отвечает спиральный вырез-катушка, изготовленный в металлических цилиндрах с внешними подвижными частями. Ключевым фактором оптимизации параметров магнита является использование новой процедуры расчета, основанной на более реалистичной модели, учитывающей геометрию магнита с нарушенной азимутальной симметрией. Целью данной статьи является теоретическое сравнение предложенной геометрии с другими существующими конструкциями. Здесь не представлен конкретный прототип. Было обнаружено, что четкое понимание характеристик катушки с вырезом является важным шагом для ее дальнейшего рассмотрения в качестве потенциальной автоадаптивной (электронно-управляемой) магнитной системы для приложений FFC.
Том. 92№. 11 стр. 1460–1463
В этой статье мы изучаем влияние гравитационных полей, создаваемых такой топологией, как пространство-время космической струны, на осциллятор Клейна-Гордона в присутствии однородного магнитного поля. Мы приводим собственные решения рассматриваемой задачи и анализируем влияние пространства-времени космических струн на собственные значения.
Том. 92№. 11 стр. 1470–1473
Исследуется взаимодействующий модифицированный газ Чаплыгина, обладающий сдвиговой и объемной вязкостью.Мы предполагаем, что объемная вязкость является функцией плотности, а также рассматриваем взаимодействие со сменой знака. Предполагается взаимодействие модифицированного газа Чаплыгина с барионным веществом. Затем исследуется влияние вязкости на космологические параметры, такие как энергия, плотность, параметр хаббловского расширения, масштабный фактор и параметр замедления. Мы можем рассматривать эту модель как игрушечную модель нашей вселенной.
Том. 92№. 11 стр. 1494–1500
Мы рассматриваем модель Вселенной с переменной G и Λ.Предметом интереса является феноменологическая модель для Λ, предложенная и рассмотренная в этой статье впервые (насколько нам известно), в предположении, что призрачная темная энергия существует и взаимодействует со Вселенной через Λ. Мы рассматриваем возможность существования необычных связей между различными компонентами флюидов во Вселенной. Мы хотели бы подчеркнуть, что это просто предположение и может быть очень далеко от реальности. Эта модель интересна феноменологически и математически, но мы не будем обсуждать физические условия или возможности реализации модификаций.
